
terinary and Animal Science, 2026
Physiological Benefits of Protected AD3E on Nutrient Digestibility, Hematology, and Small Intestinal Morphology and Carcass Characteristics in Fattening Lambs
Danyal Zarrin Kelka, Mohammad Ebrahim Nooriyan Soroorb, Fardin Hozhabric**, Hadi
Cheraghid, Khoda Bakhsh Rashidie
فواید فیزیولوژیکی AD3E محافظتشده بر قابلیت هضم مواد مغذی، پارامترهای خون، ریختشناسی روده کوچک و ویژگیهای لاشه در برههای پرواری
چکیده
مصرف کافی ویتامینهای محلول در چربی، از جمله ویتامین A، D3 و E، به دلیل نقش آنها در صلاحیت ایمنی، تنظیم متابولیک و محافظت آنتیاکسیدانی، برای برههای پرواری ضروری است. جیرههای غذایی مرسوم اغلب سطوح متناقضی از این ویتامینها را فراهم میکنند، که مکملسازی را ضروری میسازد.
مطالعه حاضر به منظور بررسی تأثیر سطوح مختلف مکمل AD3E محافظتشده با شکمبه بر عملکرد، تخمیر شکمبهای، قابلیت هضم خوراک، ویژگیهای کمی لاشه و ریختشناسی روده کوچک برههای پرواری انجام شد. بیست و چهار بره نر (۴ ماهه) با وزن بدن ۳۴.۱۱ ± ۲.۲ کیلوگرم به مدت ۷۰ روز در قالب طرح کاملاً تصادفی به سه گروه تقسیم شدند. تیمارهای آزمایشی شامل یک گروه کنترل که با جیره پایه (بدون مکمل ویتامینی) تغذیه میشدند، گروه دوم که با جیره پایه به همراه ۱ کیلوگرم در تن ویتامین AD3E تغذیه میشدند و گروه سوم که با جیره پایه به همراه ۲ کیلوگرم در تن ویتامین AD3E تغذیه میشدند، بودند. عملکرد رشد، قابلیت هضم مواد مغذی، پارامترهای تخمیر شکمبه و ویژگیهای لاشه ارزیابی شدند.
نتایج نشان داد که استفاده از AD3E محافظتشده، افزایش وزن روزانه و ضریب تبدیل غذایی برههای نر را در دو گروه دریافتکننده ویتامین در مقایسه با گروه کنترل بهبود بخشید (P < 0.05). وزن نهایی برههایی که مکمل ویتامین محافظتشده دریافت میکردند، بیشتر از گروه کنترل بود (P < 0.05). قابلیت هضم مواد مغذی بدون تغییر باقی ماند. در مقایسه با گروه کنترل، پروتوزوآهای شکمبه (جمعیت کل و انتودینیوم) در برههایی که ویتامین را در سطح 1 کیلوگرم در تن دریافت کردند، به طور قابل توجهی افزایش یافت (P < 0.05). پارامترهای خون از جمله کاهش گلوکز در 1 کیلوگرم در تن، افزایش پروتئین کل و کاهش کلسترول در 2 کیلوگرم در تن، و افزایش هماتوکریت و هموگلوبین با مکمل تغییراتی را نشان دادند. مورفولوژی روده کوچک هیچ تغییر قابل توجهی در ارتفاع پرزها یا عمق کریپت نشان نداد. صفات لاشه تحت تأثیر قرار نگرفتند.
به طور کلی، نتایج نشان داد که استفاده از مکمل ویتامین AD3E محافظتشده در جیره برهها، عملکرد برههای پرواری (وزن نهایی بدن (کیلوگرم)، افزایش وزن کل (کیلوگرم) و افزایش وزن روزانه (گرم)) را از طریق تأثیر مثبت بر تخمیر شکمبه بهبود بخشید، اگرچه بر قابلیت هضم جیره تأثیری نداشت.
کلمات کلیدی: برهها، هیستومورفومتری روده، جمعیت پروتوزوآ، تخمیر شکمبه، ویتامین، افزایش وزن.
جهت دانلود مقاله بر روی لینک زیر کلیک نمایید:
- مقدمه
برههای پرواری برای عملکرد و سلامت مطلوب به ویتامینها نیاز دارند و هر ویتامین عملکرد منحصر به فرد و غیرقابل جایگزینی دارد (Paulo et al., 2022). با توجه به تغییرات گسترده در محتوای ویتامینهای A، D3 و E در خوراکهای معمول دام، به ویژه علوفههای برداشت شده، و عواملی که بر استفاده و فراهمی زیستی آنها تأثیر میگذارند، حیوانات نمیتوانند این ویتامینهای محلول در چربی را سنتز کنند (NRC, 2021). این ویتامینها مستعد تخریب میکروبی شکمبه هستند و فراهمی زیستی را کاهش میدهند. بنابراین، اشکال محافظتشده (به عنوان مثال، کپسوله شده برای عبور از شکمبه) جذب و اثربخشی پس از شکمبه را تضمین میکنند (Sarkar et al., 2022).
این تخریب، مکملهای محافظتشده را برای تأمین نیازهای فیزیولوژیکی و حفظ تولید بالا ضروری میکند (Sarkar et al., 2022)، زیرا ویتامینهای محلول در آب و ویتامین K به اندازه کافی توسط میکروبهای شکمبه سنتز میشوند (Krogstad et al., 2025). طبق توصیههای NRC، برههای پرواری به ازای هر ۱۰۰ کیلوگرم وزن بدن روزانه به ۳۰۰۰ واحد بینالمللی رتینول، ۳۰۰ واحد بینالمللی توکوفرول و ۵۵۵ واحد بینالمللی کوله کلسیفرول نیاز دارند (NRC, 2007).
ویتامین E که برای گوسفند ضروری است اما در بافتها ذخیره نمیشود، به ویژه برای برههای جوان با ۱۰ تا ۶۰ میلیگرم در کیلوگرم ماده خشک بسیار مهم است (NRC, 1985). این ویتامینها در مجموع از سلامت حیوان حمایت میکنند (McDonald et al., 2010). خوراکهای استاندارد برای برههای پرواری اغلب فاقد ویتامینهای A، D3 و E کافی هستند و به دلیل هزینههای بالا، استفاده از یونجه را محدود میکنند. بنابراین، افزودن یونجه به جیره میتواند سلامت و رشد را افزایش دهد (McDonald et al., 2010).
ویتامین A سلولهای اپیتلیال، عملکرد اندامها و رشد را حفظ میکند (Song et al., 2023; Tanumihardjo, 2011). ویتامین D3 به تشکیل استخوان، کلسیفیکاسیون (تجمع کلسیم) و جذب کلسیم، فسفر، آهن، روی و منیزیم کمک میکند (Al-Asadi et al., 2020). ویتامین E، یک آنتیاکسیدان طبیعی، رشد، تولید مثل و پاسخ ایمنی را تقویت میکند و اکسیداسیون فسفولیپیدهای دیواره سلولی را در گاوهای شیری (Xiao et al., 2021) و دامها (Shastak et al., 2023) کاهش میدهد. مطالعات روی گوسالههای نر (Gorocica-Buenfil et al., 2007; Knutson et al., 2020) و روی گوسالهها (Scapol et al., 2023) نشان میدهد که مکمل ویتامین A حتی در سطوح پایین، هیچ تأثیر معنیداری بر افزایش وزن روزانه، مصرف ماده خشک یا ضریب تبدیل غذایی ندارد. ویتامینها بر متابولیسم کربوهیدرات و چربی تأثیر میگذارند و به طور بالقوه از طریق کاهش گلوکز خون و افزایش اشتها، مصرف خوراک را افزایش میدهند، در حالی که کمبود آنها ممکن است مصرف را کاهش دهد (McDowell et al., 2008).
ویتامینهای B، C و D تجزیه ماده خشک، متابولیسم شکمبه و تخمیر را افزایش میدهند (Witariadi et al., 2022). ویتامین E کل اسیدهای چرب فرار، تولید استات، نسبت استات به پروپیونات را افزایش میدهد و بوتیرات را در مایع شکمبه بز در شرایط آزمایشگاهی کاهش میدهد (Hou et al., 2013). در گاوهای شیری، AD3E سیستمهای ایمنی و آنتیاکسیدانی را بهبود میبخشد و GPx و SOD را افزایش میدهد، بدون اینکه بر صفات لاشه تأثیر بگذارد (Wang et al., 2022a). استفاده از ویتامین D3 (7.5×106 واحد بینالمللی) (Baldin et al. 2013) و 2×106 واحد بینالمللی (Lobo-jr et al., 2012) هیچ تاثیری بر عملکرد رشد، ویژگیهای لاشه و کیفیت گوشت در گاوهای پرواری نشان نداده است. مطالعات Ramos et al. (2024) روی چهار گروه از گوسالههای پرواری (گروه کنترل، ویتامین AD3E، ویتامین B و AD3E+B؛ نوع معمولی) نشان داده است که مکملسازی با ویتامین AD3E معمولی در جیره گوسالههای پرواری، در مقایسه با سایر گروهها، هیچ تاثیری بر pH شکمبه نداشته اما چربی درون عضلانی (ماربلینگ) را افزایش داده است.
سطوح مکمل به گونهای طراحی شده بودند که فرمهای محافظتشده را تقریباً 2.4 برابر بیشتر از نیازهای پایه NRC (2007) برای جبران کمبودهای احتمالی در شکمبه، بدون ایجاد هیپرویتامینوز، فراهم کنند، همانطور که توسط مطالعات اخیر در مورد ویتامینهای محافظتشده در شکمبه در نشخوارکنندگان پشتیبانی میشود (Shastak et al., 2023).
از آنجا که ویتامینها به تجزیه شکمبه بسیار حساس هستند، پوشینهدار میشوند (Santschi et al., 2005). با وجود نقشهای شناختهشده ویتامینهای A، D3 و E، شکافهایی در تأثیر فرمهای محافظتشده بر تحویل محافظتشده در شکمبه برههای پرواری برای افزایش فراهمی زیستی در میان رژیمهای غذایی با کنسانتره بالا وجود دارد. این مطالعه با ارزیابی 1 و 2 کیلوگرم در تن AD3E محافظتشده بر رشد، قابلیت هضم، تخمیر شکمبه، پروفایل خون، مورفولوژی روده و صفات لاشه، به این موضوع میپردازد. AD3E محافظتشده برای غلبه بر تجزیه شکمبه، بهبود راندمان مواد مغذی و تعادل میکروبی در جیره با کنسانتره بالا، با کاربردهای بالقوه در تغذیه بره، حیاتی است.
نقاط قوت این مطالعه شامل ارزیابی جامع پارامترهای فیزیولوژیکی در برههای مهربان است که شکافهای دانش در مورد اثرات اشکال محافظتشده برای استراتژیهای عملی تغذیه نشخوارکنندگان را پر میکند. بنابراین، ما فرض کردیم که مکمل ویتامینهای محلول در چربی (AD3E)، عملکرد بره، تخمیر شکمبه و ویژگیهای لاشه را بهبود میبخشد.
از آنجایی که اطلاعات کمی در مورد اثرات AD3E محافظتشده (محافظتشده با پوشش چربی شکمبه) در برههای پرواری وجود دارد، این تحقیق بررسی مزایای وابسته به دوز بر عملکرد، قابلیت هضم، تخمیر و ویژگیهای لاشه را توجیه میکند.
- مواد و روش
این آزمایشها مطابق با دستورالعملهای تعیینشدهی رفاه حیوانات انجام شد و تمام پروتکلهای آزمایشی توسط کمیته اخلاق دانشگاه رازی، کرمانشاه، ایران (شماره تأیید اخلاق: IR.RAZI.REC.1402.052) رعایت و تأیید شد. پس از انجام آزمایشها و ارائه گزارش کامل به همراه جزئیات روشهای آزمایش به کمیته مربوطه، کمیته با شماره گواهی IR.RAZI.REC.1402.063 تأیید کرد که مطالعه با تمام مقررات مطابقت دارد.
2-1 حیوانات و تیمارها
در مجموع ۲۴ بره نر چهار ماهه مهربان با میانگین وزن اولیه ۳۴.۱۱±۲.۲ کیلوگرم (انحراف معیار پایین، CV <7% برای به حداقل رساندن تغییرپذیری)، به صورت انفرادی به مدت ۷۰ روز در قالب طرح کاملاً تصادفی در ۳ گروه با ۸ تکرار نگهداری شدند. در طول دوره سازگاری (۱۴ روز)، تمام عملیات بهداشتی مانند واکسیناسیون، کنترل انگلهای داخلی و خارجی و سازگاری با تغذیه انجام شد. بر اساس NRC (2007) و برای افزایش وزن روزانه ۳۰۰ گرم، جیره غذایی گوسفندان با ترکیبی از ۳۰٪ علوفه و ۷۰٪ کنسانتره، شامل ۱۴٪ پروتئین تنظیم شد (جدول ۱).
جیره خوراکی روزانه هر حیوان در حد اشتها بود و غذای روزانه مورد نیاز گوسفندان در ساعات ۸:۰۰، ۱۴:۰۰ و ۱۹:۰۰ توزیع میشد. آب شیرین در تمام طول دوره به طور مداوم در دسترس هر بره بود.
تیمارهای آزمایشی شامل تیمار اول (کنترل): جیره پایه، بدون مکمل ویتامینی؛ تیمار دوم: جیره پایه + ویتامین AD3E مخلوط در خوراک به میزان یک کیلوگرم در تن؛ و تیمار سوم: جیره پایه + ویتامین AD3E مخلوط در خوراک به میزان دو کیلوگرم در تن. مکمل AD3E محافظتشده (محافظتشده در شکمبه با پوشش چربی) شامل 3.500.000 واحد بینالمللی در کیلوگرم، 1.200.000 واحد بینالمللی در کیلوگرم، 25 واحد بینالمللی در کیلوگرم به ترتیب ویتامین A، D3 و E و 300 میلیگرم سلنیوم در کیلوگرم بود (بهدام رشد خراسان، ایران).
در مطالعه قبلی (Nazari Darabkhani et al., 2024)، دوزهای مصرفی توصیهشده بر اساس NRC هیچ تأثیری بر رشد برههای پرواری نداشت. از سوی دیگر، نیاز دقیق برههای پرواری به ویتامین AD3E مشخص نیست؛ بنابراین، این مقادیر انتخاب شدهاند. برای تعیین پروتئین خام و خاکستر خشک از روشهای شرح داده شده در AOAC International (2002) استفاده شد. فیبر نامحلول در شوینده خنثی (NDF) و فیبر نامحلول در شوینده اسیدی (ADF) با استفاده از پروتکل Van Soest et al. (1991) تعیین شدند. انرژی قابل متابولیسم بر اساس ترکیبات جیره و دادههای جداول NRC 2007 محاسبه شد (جدول 1).
2-2 عملکرد
خوراک روزانه طبق توصیه NRC (2007) تنظیم و در آخور حیوانات قرار داده شد به طوری که 5٪ از خوراک به عنوان باقیمانده در آخور هر حیوان (n=24) باقی بماند. هر روز صبح قبل از دادن وعده غذایی صبح، باقیمانده خوراک هر بره از آخور جمع آوری و سپس خوراک تازه جایگزین شد. برای محاسبه مصرف خوراک، میزان خوراک داده شده و باقیمانده خوراک روزانه هر بره در کل دوره آزمایش ثبت شد. میزان مصرف ماده خشک (DMI) هر بره بر اساس تفریق باقیمانده خوراک روزانه هر حیوان از ماده خشک روزانه داده شده تعیین شد.
برای تخمین پارامترهای عملکرد هر بره، حیوانات به صورت هفتگی پس از 8 ساعت گرسنگی و قبل از دادن وعده غذایی صبح وزن کشی شدند. در کل دوره آزمایش، افزایش وزن کل هر بره با تفریق وزن نهایی از وزن اولیه به دست آمد. افزایش وزن روزانه هر بره نیز با تقسیم کل افزایش وزن بر تعداد روزهای پرورش (70 روز) تعیین شد. ضریب تبدیل غذایی (FCR) با تقسیم کل DMI هر بره (کیلوگرم) بر کل افزایش وزن (کیلوگرم) در طول دوره پرواربندی به دست آمد. همچنین، وزن بدن کاهش یافته، DMI مورد انتظار، DMI Obs/DMI Exp و NEm, NEg مورد انتظار طبق روش Urías-Estrada و همکاران (2023) محاسبه شدند (جدول 2).
3-2 قابلیت هضم مواد مغذی
در پایان دوره آزمایش، پنج بره (وزن بدن، ۵۳.۷۵ ± ۱.۴۳) از هر تیمار انتخاب و به مدت هفت روز در قفسهای متابولیکی قرار داده شدند (جدول ۳).
توزیع خوراک و جمعآوری مدفوع هر روز در یک زمان مشخص انجام شد. میزان خوراک مصرفی بر اساس میزان خوراک داده شده و مقدار باقیمانده در روز بعد محاسبه شد. میزان خوراک ارائه شده به حیوانات در آزمایش قابلیت هضم بر اساس نیازهای نگهداری حیوانات بود. مدفوع هر حیوان به مدت ۷ روز جمعآوری شد. تقریباً ۱۰۰ گرم نمونه مدفوع از هر حیوان هر روز برای تعیین ماده خشک گرفته شد و یک نمونه بر اساس درصد مدفوع روزانه برای آزمایشهای بعدی گرفته شد و در دمای 20- درجه سانتیگراد نگهداری شد. نمونهها از خوراک مصرفی و ضایعات خوراک روزانه برای تعیین ترکیب شیمیایی گرفته شدند (AOAC، 2002).
نمونههای مدفوع هر حیوان ذوب، همگن و برای تجزیه و تحلیل نمونهبرداری شدند. کل نمونههای مدفوع به طور مشابه جمعآوری و یک نمونه نماینده از کل نمونهها نگه داشته شد.
2-4 نمونهگیری و اندازهگیری خون
در پایان دوره آزمایش، ساعت 8:00 صبح، نمونههای خون از ورید وداج هر 8 بره/تیمار (n = 24) در لولههای هپارینه (پلاسما) و غیر هپارینه (سرم) گرفته شد؛ سپس سانتریفیوژ (3000 دور در دقیقه، 10 دقیقه) انجام شد و سرم و پلاسما در دمای 20- درجه سانتیگراد تا زمان تجزیه و تحلیل شامل گلوکز، پروتئین کل، آلبومین، کراتینین، تری گلیسیرید، کلسترول، HDL، LDL، ALP، ALT، AST، Hct، Hb، RBC، MCV، MCHC و Plt، با استفاده از دستگاه شمارش سلولی کاملاً اتوماتیک (Boule Medical، Exigo Vet، سوئد) و دستگاه اتوآنالیزر (Alpha-classic، ایران) و کیتهای تجاری (Pars Azmoon®، تهران، ایران) نگهداری شدند.
2-5 پارامترهای تخمیر
مایع شکمبه هر بره سه ساعت پس از اولین وعده غذایی (12:30 بعد از ظهر) و در پایان دوره توسط لوله مری جمعآوری شد. برای عادت دادن برهها به فرآیند جمعآوری مایع شکمبه و حذف بزاق از مایع جمعآوریشده، مایع شکمبه هر روز به مدت سه روز قبل از آزمایش از برهها جمعآوری شد و در روز چهارم، نمونه نهایی مایع شکمبه برای آزمایش جمعآوری شد. pH مایع شکمبه توسط pH متر (Hanna USA Hl98103) ثبت شد، نیتروژن آمونیاکی طبق روش Broderick & Kang (1980) با استفاده از اسپکتروفتومتر (Scan UV Visible, CARY100, VARIAN) در طول موج ۶۳۰ نانومتر اندازهگیری شد و اسیدهای چرب فرار طبق روش Ottenstein & Bartley (1971) با استفاده از GC (Unicam 4410, UK) اندازهگیری شدند. از ۴-متیل والریک اسید (سیگما) به عنوان استاندارد داخلی استفاده شد. میزان اتلاف انرژی به شکل گاز متان تولید شده بر اساس روابط زیر تخمین زده شد (Klasing 2007).
معادله 1)
CH4 (MJ/d) = 3.96 (±1.18) + 0.561(±0.130) × DMI (kg/d)
معادله 2)
CH4 (MJ/d) = 2.70 (±1.38) + 1.16(±0.271) × DMI (kg/d) – 15.8(±6.86) × EE (kg/d)
معادله 1 بر اساس ضریب ثابت و DMI است و در معادله 2، ضریب ثابت، DMI و چربی خام خوراک و میزان گاز متان تولید شده بر حسب مگاژول در روز تخمین زده شد.
2-6 شمارش پروتوزآها
شناسایی و شمارش پروتوزآها با استفاده از نرمافزار Dinocapture نصبشده روی کامپیوتر، میکروسکوپ نوری و لامل هموسیتومتر طبق روش توصیهشده (Dehority 2003) انجام شد. لامل هموسیتومتر با آب مقطر شسته و سپس با یک حوله کاغذی نرم خشک شد. سپس، محتویات یک لوله فالکون 50 میلیلیتری به آرامی مخلوط شد. با استفاده از سرنگ، یک قطره از نمونه به حفره مرکزی لامل هموسیتومتر منتقل شد، لامل روی لامل شمارش قرار داده شد و شمارش با بزرگنمایی 10 برابر با استفاده از میکروسکوپ انجام شد.
جنس پروتوزآها بر اساس تعداد نواحی مژکدار و توزیع مژکها روی سلول و غلظت کل آنها در هر میلیلیتر از محتویات شکمبه محاسبه شد.
2-7 ویژگیهای لاشه، نمونهبرداری و مورفومتری روده کوچک
سه بره به طور تصادفی (انتخاب تصادفی تضمین شده بود، از تعصب جلوگیری شد) در هر تیمار مستقل از زیرمجموعه قابلیت هضم (n = 8) انتخاب شدند، وزنکشی شده و در پایان آزمایش ذبح شدند تا ویژگیهای لاشه و وزن اندامها شامل وزن پس از کشتار، وزن لاشه، وزن پشم و پوست، طول لاشه، عرض لاشه، طول ران و طول دست طبق روش Asadian et al. (1996) بررسی شود. علاوه بر این، روده کوچک برهها سپس از طریق تشریح (جدول 8) برداشته شد. سپس، نمونهای تقریباً 5 سانتیمتری از دوازدهه، ژژنوم و ایلئوم گرفته شد.
بافتها به مدت چهار روز در فرمالین 10٪ تثبیت شدند، رنگآمیزی شدند و طبق روشهای استاندارد در پارافین قرار گرفتند (یون و همکاران، 2013). برای گرفتن عکس از میکروسکوپ نوری استفاده شد.
از نرمافزار تصویربرداری برای اندازهگیری طول پاپیلا، عرض پاپیلا و عمق کریپت در دوازدهه، ژژنوم و ایلئوم استفاده شد (Axiovision، 8.4؛ Carl Zeiss، Oberkochen، آلمان).
2-8 تجزیه و تحلیل آماری
این آزمایش در قالب طرح کاملاً تصادفی با سه تیمار و هشت تکرار (بره) در هر تیمار انجام شد. دادههای بهدستآمده برای هر صفت با استفاده از نرمافزار SPSS (نسخه 27) در سطح (α = 0.05) و مدل آماری زیر تجزیه و تحلیل شدند. آزمون چند دامنهای دانکن برای مقایسههای پس از تجزیه و تحلیل واریانس (ANOVA) استفاده شد، زیرا برای گروههای با اندازه برابر و کنترل خطای نوع اول مناسب است (Montgomery ، 2017). نرمال بودن و همگنی دادهها از طریق آزمونهای شاپیرو-ویلک (P>0.05) و لون (P>0.05) تأیید شد. نیازی به تبدیل نبود.
مدل آماری به شرح زیر بود:
yij= μ + Ti + eij
که در آن: yij = i امین مشاهده از j امین تیمار، μ = میانگین کل، Ti = i امین تیمار و eij = اثرات باقیمانده.
- نتایج و بحث
3-1 عملکرد
جدول ۲ نشان میدهد که تجویز ویتامین AD3E محافظتشده به طور قابل توجهی بر معیارهای مختلف عملکرد در برههای پرواری در طول دوره آزمایش تأثیر گذاشت. وزنهای اولیه در تمام تیمارها قابل مقایسه بودند که یکنواختی را در شرایط پایه تأیید میکند. با این حال، وزن نهایی بدن (FBW) به طور قابل توجهی در تیمارهای ۲ و ۳ در مقایسه با گروه کنترل بالاتر بود، که نشان میدهد مکمل ویتامین رشد بیشتری را تسهیل کرده است. افزایش وزن کل از الگوی مشابهی پیروی کرد، به طوری که تیمار ۲ بیشترین افزایش وزن را ثبت کرد که به طور قابل توجهی بیشتر از گروه کنترل بود، در حالی که تیمار ۳ پاسخ متوسطی را نشان داد.
علاوه بر این، میانگین افزایش وزن روزانه (ADG) در تیمار ۲ نسبت به گروه کنترل به طور قابل توجهی بهبود یافت و تیمار ۳ دوباره نتایج متوسطی را نشان داد. بنابراین، افزودن AD3E به جیره غذایی به میزان ۱ کیلوگرم در تن منجر به افزایش قابل توجه FBW و ADG (P<0.05) شد، بدون اینکه هیچ اثر قابل تشخیصی بر DMI داشته باشد.
بهبود ۱۶.۵ درصدی در افزایش وزن روزانه که با دوز متوسط ویتامین مشاهده شد، نشاندهنده افزایش راندمان خوراک است. با افزودن ۱ کیلوگرم در تنAD3E (P = 0.03)، بهبود ۱۲ درصدی در ضریب تبدیل خوراک (FCR) مشاهده شد که نشاندهنده استفاده کارآمدتر از مواد مغذی است.
این یافته از نقش تثبیتشده ویتامینها در افزایش راندمان متابولیک بدون افزایش مصرف خوراک پشتیبانی میکند (McDowell, 2008). بنابراین، ویتامینها ممکن است نقش مهمی در متابولیسم کربوهیدراتها، چربیها، پروتئینها و همچنین رشد حیوان داشته باشند (McDowell et al., 2008).
جدول 2
مطالعات محدودی مکمل AD3E را بر عملکرد پرواربندی حیوانات بررسی کردهاند و اکثر مطالعات ویتامینها را به صورت جداگانه بررسی کرده و نتایج متناقضی را گزارش کردهاند. ویتامین AD3E افزایش وزن گوساله را در مقایسه با گروه کنترل بهبود بخشید و بیان شد که این بهبود در گروه مصرفکننده ویتامین AD3E میتواند مربوط به تأثیر ویتامینهای A، D3 و E بر افزایش اشتها و خوشطعمی جیره باشد که باعث افزایش مصرف ماده خشک و در نهایت بهبود رشد و عملکرد حیوانات تحت آزمایش شده است (Habeeb et al., 2018).
این محققان همچنین اظهار داشتند که بهبود قابل توجه در افزایش وزن گوساله به دلیل مکمل ویتامین A و E ممکن است به این دلیل باشد که این ویتامینها در سنتز کوآنزیمهای مهم (NAD و FAD) که مسئول فرآیندهای اکسیداتیو بیولوژیکی هستند که ATP مورد نیاز برای بیوسنتز پروتئینها، چربیها و کربوهیدراتها را تولید میکنند، نقش دارند. مشابه یافتههای مطالعه حاضر، Al_Galiby (2022) نیز در سه گروه از بزغالههای در حال رشد نشان داد که تجویز عضلانی ویتامین AD3E دو بار در هفته به طور قابل توجهی افزایش وزن روزانه و کل افزایش وزن بزغالههای در حال رشد را بهبود میبخشد. مکمل ویتامین E در برههای Karya هیچ تأثیر معنیداری بر افزایش وزن روزانه نشان نداد، اما ضریب تبدیل غذایی را حدود 10٪ بهبود بخشید، که نشان میدهد ویتامین E ممکن است راندمان متابولیکی را در نشخوارکنندگان پرواری افزایش دهد (Atay et al., 2009).
بهبود معیارهای رشد و FCR مشاهده شده با مکمل AD3E محافظتشده را میتوان به سه مسیر فیزیولوژیکی متمایز اما همافزایی نسبت داد: (1) تنظیم غدد درونریز محور سوماتوتروپیک (ویتامین A)، (2) افزایش کانیسازی اسکلتی و استفاده از مواد مغذی (ویتامین D3) و (3) فعالیت آنتیاکسیدانی محافظتکننده سلولی با اثرات ثانویه ایمونومتابولیک (ویتامین E).
واضحترین مکانیسم شامل ویتامین A (رتینول) است که به عنوان یک تنظیمکننده تغذیهای مستقیم محور سوماتوتروپیک عمل میکند. Oka et al. (2004) نشان دادند که حیواناتی که مکمل کافی ویتامین A دریافت میکنند، غلظت IGF 1 سرمی به طور قابل توجهی بالاتر و ADG بالاتری در مقایسه با حیواناتی که ویتامین A دریافت نمیکنند، نشان میدهند، علیرغم DMI معادل و عدم تفاوت قابل تشخیص در هورمون رشد (GH)؛ با این حال، نشان داده شده است که اسید رتینوئیک، نوعی از ویتامین A، بیان ژن هورمون رشد را نیز تنظیم میکند (Cheung et al., 2020). این نشان میدهد که ویتامین A اثر تقویتکننده رشد خود را نه با افزایش ترشح GH، بلکه با افزایش سنتز و/یا ترشح IGF-1 کبدی اعمال میکند.
این یافتهها با کارهای مولکولی قبلی در مدلهای پرندگان سازگار است که نشان میدهد کمبود ویتامین A بیان ژن IGF-1 را در بافت کبدی کاهش میدهد (Fu et al., 2001). با توجه به اینکه IGF-1 واسطه اصلی رشد سوماتیک وابسته به GH و احتباس نیتروژن است، افزایش IGF-1 در گردش خون، یک مکانیسم غدد درون ریز مستقیم برای بهبود افزایش وزن و راندمان خوراک مشاهده شده در مطالعه ما فراهم میکند.
ویتامین D3 (کوله کلسیفرول) از طریق عملکرد کلاسیک خود در هموستاز کلسیم و فسفر به عملکرد رشد کمک میکند. مطالعات مربوط به مکمل AD3E تأیید میکند که ویتامین D3 راندمان جذب کلسیم و فسفر روده را افزایش میدهد، که برای تشکیل استخوان و کلسیفیکاسیون ضروری است (Nazari Darabkhani et al., 2024). در برههای پرواری با رشد سریع، معدنی شدن کافی اسکلت یک عامل مجاز برای افزایش بافت بدون چربی است.
فراتر از متابولیسم مواد معدنی، شواهد نوظهور در تغذیه نشخوارکنندگان نشان میدهد که وضعیت ویتامین D بر بیان ژن مربوط به رشد عضلات و ترکیب لاشه تأثیر میگذارد (Hernández and Tamassia, 2025). بنابراین، بهبود ضریب تبدیل غذایی ممکن است نشاندهنده کاهش هزینه متابولیکی حفظ اسکلت باشد و امکان تقسیم بخش بیشتری از انرژی غذایی به سمت سنتز پروتئین ماهیچه را فراهم کند. ویتامین E با خنثی کردن گونههای فعال اکسیژن تولید شده در طول جریان متابولیکی بالا، سیالیت و عملکرد غشا را حفظ میکند. این اثر محافظت سلولی به ویژه در برههای با رشد سریع، که در آن استرس اکسیداتیو میتواند عملکرد میوسیتها و هپاتوسیتها را مختل کند، اهمیت دارد (Shojadoost et al., 2021). علاوه بر این، ویتامین E اثرات تعدیلکننده سیستم ایمنی را اعمال میکند که به طور غیرمستقیم از رشد پشتیبانی میکند.
ویتامین E با بیان سیتوکینیهای پیشالتهابی (مانند IL 1b، IL-6) و افزایش پاسخهای لنفوسیتهای T (CD4+، CD8+) و لنفوسیتهای B، انحراف مواد مرتبط با سیستم فعالسازی ایمنی بالینی را کاهش میدهد (Shojadoost et al., 2021). کاهش BUN سرم و تغییر نیتروژنیکی مشاهده شده در تیمارهای AD3E محافظت شده با شکمبه (Nazari Darabkhani et al., 2024) ممکن است بهبود نیتروژن و تقسیم بندی کارآمدتر نیتروژن در شکم به دلیل کاهش کاتابولیسم اسیدهای آمینه سازگار باشد.
برخی از این مراکز خاص به طور مستقیم در مطالعه حاضر گزارش نشده اند، اما از نظر فیزیولوژیکی با بهبودهای مشاهده شده 12 تا 16.5 درصد در افزایش وزن و ضریب تبدیل مطابقت دارند. بهبود ضریب تبدیل غذایی بدون افزایش همزمان در DMI قویاً نشان میدهد که افزایش عملکرد مشاهدهشده به دلیل افزایش راندمان متابولیکی است تا مصرف مصرف را افزایش دهد.
ترکیبی از حرکت غدد درونریز (آنابولیسم با مصرف کننده IGF-1)، بهبود یافتن مواد معدنی اسکلتی و کاهش هزینههای زیست محیطی منسجمی، توضیح فیزیولوژیکی را برای این موضوع ارائه میدهد که چرا مکمل AD3E محافظت میکند، برهها را میسازد تا هر واحدی را از دست بدهند.
3-2 قابلیت هضم
جدول 3 هیچ اثر معنیداری از مکمل ویتامین AD3E محافظتشده بر قابلیت هضم مواد مغذی، به جز قابلیت هضم پروتئین، نشان نمیدهد. به طور مشابه، Kannan et al. (2024) نشان دادند که مکمل 25-هیدروکسی ویتامین D3 بر قابلیت هضم مواد مغذی در بزها تأثیری ندارد.
افزایش وابسته به دوز در قابلیت هضم پروتئین خام مشاهده شده با مکمل AD3E در سطح 2 کیلوگرم در تن، اما نه در سطح 1 کیلوگرم در تن، را میتوان به چندین مکانیسم فیزیولوژیکی و متابولیکی مرتبط نسبت داد، اگرچه شواهد مستقیم برای پاسخهای قابلیت هضم پروتئین به طور خاص در مقالات محدود است.
جدول 3
ویتامین A برای حفظ یکپارچگی بافت اپیتلیال در سراسر دستگاه گوارش ضروری است و ویتامین A و ویتامین D بیان پروتئینهای اتصال محکم روی سلولهای اپیتلیال روده را که برای عملکرد سد در روده حیاتی هستند، تنظیم میکنند (Cantorna et al., 2019). ویتامین A نقش اساسی در بینایی و تمایز سلولی دارد، که مورد دوم یک مکانیسم اصلی منحصر به فرد را ارائه میدهد که به توضیح تأثیر ویتامین A بر موانع اپیتلیال کمک میکند و تغییرات در پوشش اپیتلیال اندامهای حیاتی در اوایل کمبود رخ میدهد، که نشان دهنده نقش بالقوه مهم برای عملکرد سد است (McCullough et al., 1999).
اسید رتینوئیک تمام ترانس (ATRA) با تنظیم بیان پروتئینهای اتصال محکم مانند zona occludens-1 (ZO-1)، claudin-1 و occludin برای حفظ یکپارچگی سد اپیتلیال روده بسیار مهم است (Skawratananond et al., 2025). وضعیت کافی ویتامین A، معماری بهینه پرزها و عملکرد انتروسیتها را تضمین میکند که مستقیماً بر ظرفیت جذب مواد مغذی تأثیر میگذارد. سطح ۲ کیلوگرم در تن ممکن است رتینول کافی برای پشتیبانی از این عملکردهای نگهداری اپیتلیال را فراهم کرده باشد، در حالی که سطح ۱ کیلوگرم در تن، اگرچه برای نیازهای پایه کافی است، اما ممکن است از آستانه مورد نیاز برای بهبود قابل اندازهگیری در قابلیت هضم پروتئین فراتر نرفته باشد. ویتامین D3، فرم فعال بیولوژیکی 1،25-دی هیدروکسی ویتامین D3، با تنظیم افزایشی ناقلهای کلسیم اپیتلیال، راندمان جذب کلسیم روده را افزایش میدهد (Reddy & Jialal, 2022 ).
این نقش در متابولیسم کلسیم، پیامدهای غیرمستقیم اما حیاتی برای هضم پروتئین دارد؛ یونهای کلسیم، کوفاکتورهای ضروری برای پایداری و فعالیت بهینه آنزیمهای پروتئولیتیک پانکراس، مانند تریپسین و کیموتریپسین، در روده کوچک هستند. علاوه بر این، کلسیم برای ترشح این آنزیمها از پانکراس مورد نیاز است. با افزایش جذب کلسیم، مکمل ویتامین AD3E با دوز بالا (2 کیلوگرم در تن) ممکن است فعالیت آنزیم پانکراس و محیط لومینال برای پروتئولیز را بهبود بخشیده و در نتیجه قابلیت هضم پروتئین خام را افزایش دهد.
Silva et al (2020) نشان دادند که مکمل ویتامین ADE در گاوها با موفقیت غلظت 25-هیدروکسی ویتامین D در گردش را افزایش داده و فراهمی زیستی ویتامینهای مکمل را تأیید میکند. اگرچه مطالعه آنها بهبودی در قابلیت هضم کلی مواد مغذی مشاهده نکرد، نویسندگان اذعان کردند که وضعیت ویتامین با موفقیت افزایش یافته است، که ممکن است شرایط مناسبی را برای عملکرد گوارشی در شرایط خاص رژیم غذایی ایجاد کند. علاوه بر این، اپیتلیوم دستگاه گوارش به سرعت در حال تغییر است و به ویژه در شرایط تولید فشرده، مستعد استرس اکسیداتیو است.
در سطح مکمل بالاتر (2 کیلوگرم در تن)، آلفا-توکوفرول ممکن است محافظت آنتیاکسیدانی کافی برای حفظ یکپارچگی انتروسیتها و فعالیت آنزیمهای حاشیه مسواکی فراهم کرده باشد ( Reddy& Jialal, 2022)، در نتیجه هضم پروتئین و جذب اسید آمینه کارآمدتر را تسهیل میکند.
3-3 پارامترهای خون
تغییرات مشاهده شده در پروفایل متابولیتهای خون پس از مکمل AD3E محافظتشده بینشی در مورد سازگاریهای متابولیکی زمینهساز بهبود عملکرد رشد در برههای پرواری ارائه میدهد. اثرات وابسته به دوز بر گلوکز، پروتئین کل و بخشهای لیپیدی، نقشهای فیزیولوژیکی تثبیتشده ویتامینهای A، D3 و E را در متابولیسم واسطهای، تجمع بافت و دفاع آنتیاکسیدانی منعکس میکند.
غلظت گلوکز پلاسما به طور قابل توجهی پایینتر (جدول 4) در برههایی که 1 کیلوگرم در تن AD3E محافظتشده (تیمار 2) دریافت میکنند در مقایسه با هر دو گروه کنترل و تیمار 2 کیلوگرم در تن (تیمار 3) بررسی اثرات واسطهگریشده توسط ویتامین بر هموستاز گلوکز را ضروری میسازد. این یافته با مشاهدات Nazari Darabkhani et al. (2024) همسو است، که گزارش دادند اگرچه غلظت گلوکز پلاسما در مطالعه آنها به طور قابل توجهی تحت تأثیر مکمل AD3E قرار نگرفت، اما سطح گلوکز در گروههای مکملشده با ویتامین از نظر عددی پایینتر بود و در طول دوره آزمایش از روز 30 تا 60 کاهش یافت.
تجویز مکمل AD3E در گاوها با کاهش غلظت گلوکز خون نسبت به گروههای بدون مکمل همراه بود (Toma et al., 2021). در مقابل، سطح گلوکز خون در گاومیشهای مدیترانهای تحت تأثیر مکمل ویتامین قرار نگرفت (Verdurico, 2012).
جدول 4
ترکیب ویتامین A که حساسیت به انسولین را بهبود میبخشد و ویتامین D که ترشح انسولین را افزایش میدهد، محیطی همافزایی برای دفع کارآمد گلوکز ایجاد میکند. شواهد اخیر از Yu et al. (2024) نشان داد که کمبود ویتامین A منجر به اختلال در ترشح انسولین تحریکشده توسط گلوکز و از دست دادن بیان پپتید-1 شبهگلوکاگون رودهای (GLP-1) میشود.
بازسازی ویتامین A این پارامترها را از طریق فعالسازی مسیر سیگنالینگ گیرنده بتای اسید رتینوئیک (RARβ) نرمال کرد. این مکانیسم رودهای نشاندهنده یک مسیر اضافی است که از طریق آن مکمل ویتامین ممکن است بر هموستاز گلوکز تأثیر بگذارد. عدم وجود این اثر هیپوگلیسمی در سطح مکمل بالاتر (2 کیلوگرم در تن) ممکن است نشاندهنده یک پدیده آستانه یا تعامل پیچیده بین وضعیت ویتامین و سایر هورمونهای متابولیک باشد. افزایش همزمان پروتئین کل در این سطح نشان میدهد که اسیدهای آمینه ممکن است به سمت گلوکونئوژنز هدایت شده باشند تا از افزایش تقاضای سنتز پروتئین پشتیبانی کنند.
غلظت بالای پروتئین کل در برههایی که ۲ کیلوگرم در تن AD3E محافظتشده (تیمار ۳) دریافت کردند، در مقایسه با هر دو گروه کنترل و ۱ کیلوگرم در تن، نشاندهنده بهبود احتباس نیتروژن و افزایش ظرفیت آنابولیک پروتئین در سطح بالاتر مکمل است که با شواهد مستقیم حاصل از مطالعات مکمل ویتامین E گوسفند مطابقت دارد (Ismail Mahmud & Karadas, 2023). این یافته به ویژه با توجه به اینکه غلظت آلبومین در طول تیمارها بدون تغییر باقی ماند، قابل توجه است، که نشان میدهد افزایش، منعکسکننده تغییرات در بخش گلوبولین یا سایر اجزای پروتئین است تا وضعیت هیدراتاسیون. غلظت پروتئین کل و گلوبولین برهها با تزریق ویتامین AD3E افزایش یافت و این افزایش در ۴ میلیلیتر به ازای هر حیوان بیشتر از سایر گروهها بود (Habeeb et al. 2015). افزایش پروتئین کل ممکن است به دلیل نقش فعال ویتامینها در حفظ فشار اسمزی سلولها، حفظ فرآیند سنتز پروتئین و آلبومین و افزایش اثربخشی آنها در سنتز پروتئینهای سلولی باشد (Habeeb et al., 2008).
Ismail Mahmud & Karadas (2023) مکمل ویتامین E (آلفا توکوفرول استات) را در برههای آواسی بررسی کردند و گزارش دادند که ویتامین E در رژیم غذایی با دوز ۲۰۰ و ۴۰۰ میلیگرم در روز به ازای هر بره، پروتئین کل سرم را به طور قابل توجهی افزایش داده و در عین حال غلظت گلوکز را کاهش میدهد. این شواهد مستقیم از تحقیقات گوسفند، یافتههای حاضر را تأیید میکند و تأیید میکند که ویتامین E در بهبود وضعیت پروتئین در برههای در حال رشد نقش دارد.
عملکرد آنتیاکسیدانی ویتامین E با حفظ ساختارهای سلولی دخیل در سنتز پروتئین، به وضعیت پروتئین کمک میکند. Njeru et al. (1995) ثابت کردند که غلظت توکوفرول سرم و کبد به طور قابل اعتمادی منعکس کننده میزان مصرف ویتامین E در گاوهای جوان است و تأیید میکند که ویتامین E مکمل شده، زیستفراهم است و در بافتهایی که میتواند اثرات محافظت سلولی داشته باشد، گنجانده میشود. این عملکرد تثبیتکننده غشا، با کاهش آسیب اکسیداتیو، از ظرفیت سنتز پروتئین پایدار در سلولهای کبدی و میوسیتها پشتیبانی میکند. ماهیت وابسته به دوز این پاسخ - که فقط در ۲ کیلوگرم در تن معنیدار است - نشان میدهد که در حالی که سطح پایینتر مکمل، نیازهای پایه را برآورده میکند، سطح بالاتر، مازاد ویتامین کافی را برای تحریک حداکثری مسیرهای آنابولیک پروتئین فراهم میکند.
کاهش غلظت کلسترول و LDL در گروههای مکمل (جدول 4) با اثرات هیپوکلسترولمی اثباتشده ویتامین E در برهها (Ismail Mahmud & Karadas, 2023) و تنظیم متابولیسم لیپید با واسطه ویتامین A (Chen, 2021) همسو است. این سازگاریهای متابولیکی، توضیحی مکانیکی برای بهبود عملکرد رشد و راندمان خوراک با مکمل AD3E محافظتشده ارائه میدهند. اثر هیپوکلسترولمی مشاهدهشده در مطالعه حاضر با یافتههای قبلی در مدلهای گوسفند سازگار است، جایی که مصرف ویتامین A با کاهش غلظت کلسترول همراه بود (Li et al., 2015).
شواهد از Śmiecińska et al. (2023) نشان داد که مصرف بیش از حد طولانی مدت ویتامین A با کاهش کلسترول پلاسما مرتبط است، پدیدهای که به تأثیر ویتامین A بالا در رژیم غذایی بر سنتز و کاتابولیسم کلسترول کبدی نسبت داده میشود. این تعدیلها در پروفایل متابولیتهای خون، ضرورت فیزیولوژیکی مکمل ویتامین AD3E را برای حفظ هموستاز سیستمیک و پشتیبانی از عملکردهای متابولیک حیاتی در حیوانات تولیدی منعکس میکند (Ferdouse et al., 2024).
اسید رتینوئیک تمایز آدیپوسیتها و هموستاز لیپید را از طریق سیگنالینگ گیرنده هستهای تنظیم میکند و بر لیپوژنز و پاکسازی لیپید تأثیر میگذارد (Chen, 2021).
کاهش کلسترول و LDL در 2 کیلوگرم بر تن احتمالاً نشاندهنده افزایش تنظیم متابولیسم لیپید با واسطه ویتامین A است که لیپیدها را از گردش خون دور کرده و به سمت اهداف تولیدی مانند سنتز غشاء در بافتهای در حال رشد هدایت میکند. اثرات کاهشدهنده لیپید همچنین ممکن است از طریق عملکرد ویتامین E به عنوان یک آنتیاکسیدان لیپوفیلیک که لیپوپروتئینهای در گردش را از تغییرات اکسیداتیو محافظت میکند، عمل کند. ویتامین E با کاهش پراکسیداسیون لیپید، پاکسازی ذرات LDL از گردش خون را افزایش داده و از تجمع آنها جلوگیری میکند (Ismail Mahmud & Karadas, 2023). آنها همچنین گزارش دادهاند که ویتامین E با دوز ۴۰۰ میلیگرم در روز به ازای هر بره در روز، غلظت کلسترول را در برههای آواسی به طور قابل توجهی به ۴۰.۵ میلیگرم در دسیلیتر در مقایسه با گروههای بدون مکمل کاهش میدهد.
این یافته مستقیماً با نتایج حاضر مطابقت دارد و تأیید میکند که ویتامین E اثرات کاهشدهنده کلسترول را در برههای در حال رشد اعمال میکند. Luan et al. (2023) نیز گزارش دادند که مکملسازی گاوهای نر در حال پروار با ۵۰۰ واحد بینالمللی در روز ویتامین E منجر به کاهش غلظت کلسترول سرم و بهبود وضعیت آنتیاکسیدانی میشود. نویسندگان این اثر را به نقش شناختهشده ویتامین E در مهار پراکسیداسیون لیپید در لیپوپروتئینهای در گردش خون نسبت دادند، در نتیجه متابولیسم لیپید را تعدیل کرده و پایداری اکسیداتیو را افزایش میدهند. این مشاهده که غلظت تری گلیسیرید تحت تأثیر مکمل ویتامین قرار نگرفته است، با یافتههای Njeru et al. (1995) سازگار است، که هیچ اثر درمانی ویتامین E را بر بخشهای چربی خون (کلسترول و تری گلیسیرید) در گاوهای یک ساله گزارش نکردند. این نشان میدهد که اثرات ویتامینها بر متابولیسم لیپید ممکن است ترجیحاً بر هموستاز کلسترول تأثیر بگذارد تا متابولیسم تریگلیسیرید.
پاسخهای متفاوت مشاهده شده بین سطوح مکمل، بینشی در مورد ماهیت وابسته به دوز اثرات ویتامین بر متابولیسم ارائه میدهد. گلوکز پایینتر در ۱ کیلوگرم بر تن، همراه با پروتئین کل بالا تنها در ۲ کیلوگرم بر تن، نشاندهنده نیازهای متمایز ویتامین برای مسیرهای متابولیکی مختلف است. هموستاز گلوکز ممکن است نسبت به افزایش متوسط وضعیت ویتامین حساستر باشد، که به طور بالقوه از طریق اثرات سریع بر سیگنالینگ انسولین و هورمونهای اینکرتین رودهای واسطه میشود (Dakshinamurti, 2015; Yu et al., 2024). در مقابل، آنابولیسم پروتئین - که نیاز به اثرات یکپارچه بر بیان ژن، حساسیت هورمونی و یکپارچگی سلولی دارد - ممکن است برای دستیابی به پیشرفتهای قابل اندازهگیری، نیاز به مصرف بیشتر ویتامین داشته باشد (Chen, 2021).
در مطالعه حاضر، فعالیتهای آلکالین فسفاتاز (ALP)، آلانین آمینوترانسفراز (ALT) و آسپارتات آمینوترانسفراز (AST) تحت تأثیر مکمل AD3E قرار نگرفتند. در مقابل، اسماعیل Ismail Mahmud & Karadas (2023) گزارش دادند که برههای تغذیهشده با ویتامین E، فعالیتهای AST و ALT کمتری را نشان دادند که نشاندهنده افزایش وضعیت عملکردی کبد است که منجر به متابولیسم کارآمدتر لیپید میشود. به طور مشابه، در مطالعهای روی بزغالهها، ویتامین E در رژیم غذایی (تا 1000 واحد بینالمللی) منجر به کاهش قابل توجه فعالیتهای AST و ALT در مقایسه با گروه کنترل شد. مستقیمترین ارتباط بین اجزای AD3E و آلکالین فسفاتاز (ALP) از طریق ویتامین D است.
تحقیقات نشان داده است که اختلالات متابولیسم ویتامین D، مانند دیپروویتامینوز، با فعالیت بالای ALP در سرم خون همراه است (Velykyĭ et al., 2010). این به این دلیل است که ویتامین D هموستاز کلسیم و فسفات را تنظیم میکند و تغییرات در وضعیت آن مستقیماً بر فعالیت استئوبلاست تأثیر میگذارد که ایزوفرم استخوانی ALP را تولید میکند.
نشان داده شده است که ویتامین E بر فعالیت آنزیمهای مسئول فعال کردن ویتامین D در کبد (ویتامین D3 25-هیدروکسیلاز) تأثیر میگذارد و در شرایط ویتامین D اضافی (D-هیپروویتامینوز)، به عادیسازی متابولیسم مواد معدنی، محتوای کلسیم، فسفاتها و فعالیت ایزوفرم آلکالین فسفاتاز در سرم خون کمک میکند (Velykyĭ et al., 2010). این نشان دهنده یک تعامل پیچیده است که در آن ویتامین E از تنظیم متابولیک پشتیبانی میکند.
عدم تغییر در ALP در آزمایش ما ممکن است نشان دهد که ویتامینهای مکملشده هیچ عارضه جانبی مانند کلستاز یا تغییر عمده و نامنظم در بازسازی استخوان ایجاد نکردهاند. سطح پایدار ALP نشان میدهد که متابولیسم مواد معدنی و استخوان در برههای مکملشده به خوبی تنظیم شده است.
کاهش شاخص MDA مشاهده شده با مکمل AD3E محافظتشده در مطالعه حاضر، شواهد روشنی از کاهش پراکسیداسیون لیپید و بهبود وضعیت آنتیاکسیدانی در برههای دریافتکننده مکمل ارائه میدهد (شکل 1). این اثر در درجه اول توسط فعالیت آنتیاکسیدانی زنجیرهشکن ویتامین E در غشاهای سلولی واسطهگری میشود (Li et al., 2015; Simitzis et al., 2019)، و با مشارکتهای همافزایی بالقوه ویتامینهای A و D در حفظ یکپارچگی سلولی و پشتیبانی از ظرفیت کلی آنتیاکسیدانی (Dimri et al., 2013; Strickland et al., 2021). کاهش استرس اکسیداتیو نشاندهنده مکانیسم مهمی است که در بهبود عملکرد رشد و راندمان خوراک مشاهده شده با مکمل AD3E نقش دارد.
شکل 1
جدول 5 افزایش قابل توجهی در هماتوکریت (Hct) و هموگلوبین (Hb) با مکمل AD3E محافظتشده نشان میدهد، که مقادیر آن در تیمارهای 2 و 3 در مقایسه با گروه کنترل بالاتر است (P < 0.05)، که نشان دهنده افزایش اریتروپوئز و ظرفیت انتقال اکسیژن است که احتمالاً توسط ویتامین D3 و E هدایت میشود (Keywanloo et al., 2021). تعداد گلبولهای قرمز، MCV، MCHC و تعداد پلاکتها هیچ تفاوت معنیداری نشان ندادند، که نشان دهنده اثرات انتخابی بر کیفیت گلبولهای قرمز به جای کمیت است (Samarin et al., 2022). این نتایج نشان میدهد که مکمل AD3E از سلامت خونشناسی در برههای پرواری بدون ایجاد پلیسیتمی یا ترومبوسیتوپنی پشتیبانی میکند.
جدول 5
شکل 2
میزان هموگلوبین با توجه به سطح تغذیه متفاوت است (Kitchenham et al., 1975) زیرا هموگلوبین با افزایش مصرف خوراک افزایش مییابد (El-Sabban et al., 1971). در مطالعه حاضر، برههایی که با رژیم غذایی حاوی AD3E تغذیه شدند، غلظت هموگلوبین بالاتری نیز داشتند. گزارش شده است که تغییرات در مصرف پروتئین یکی از عوامل مؤثر بر ترکیب خون است، زیرا هموگلوبین به تغییرات در مصرف پروتئین پاسخ میدهد (Treacher, 1978).
افزایش Hct و Hb در گروههای مکملشده با مطالعهای در سال ۲۰۲۱ روی گاوهای نر هلشتاین مطابقت دارد، که در آن ویتامین D3 تزریقی (۲۲۰۰۰ واحد بینالمللی در کیلوگرم) به طور قابل توجهی هموگلوبین و هماتوکریت را به صورت وابسته به دوز افزایش داد، که به نقش ویتامین D در بلوغ گلبولهای قرمز نسبت داده میشود (Keywanloo et al., 2021).
به طور مشابه، یک مطالعه در سال ۲۰۲۲ روی برههای در حال رشد با مواد معدنی کمیاب آلی (از جمله همافزایی با ویتامینها) تعداد گلبولهای قرمز و مقاومت اسمزی بالاتری را گزارش کرد که از بهبود هماتولوژی مشابه اثرات AD3E در اینجا پشتیبانی میکند (Samarin et al., 2022).
در مقابل، مطالعهای روی خوکها نشان داد که 25(OH)D3 ایمنی را از طریق افزایش ایمونوگلوبولینها افزایش میدهد اما تغییرات هماتولوژیکی را به تفصیل شرح نداد، اگرچه به مزایای سیستمیک گستردهتری اشاره داشت که میتواند به طور غیرمستقیم از پارامترهای خون پشتیبانی کند (Upadhaya et al., 2023). گزارش شده است که کمبود ویتامین A باعث کاهش تولید لنفوسیتهای T و B، اختلال در فاگوسیتوز، افزایش عفونتها و در نتیجه کاهش عملکرد مثبت سیستم ایمنی میشود (Klasing 2007). مکمل محافظتشده AD3E شامل 300 میلیگرم سلنیوم در کیلوگرم بود که با ویتامین E به عنوان یک آنتیاکسیدان برای افزایش عملکرد سیستم ایمنی و کاهش استرس اکسیداتیو در برههای پرواری همافزایی داشت (Giadinis et al., 2000).
گنجاندن سلنیوم احتمالاً از طریق همافزایی آنتیاکسیدانی با ویتامین E به بهبود جمعیت پروتوزوآ و هماتولوژی کمک کرده است (Novelli et al., 2023). گزارش شده است که سلنیوم و ویتامین E در گوسفند باعث تولید آنتیبادی شده و سطح مقاومت در دامها را به میزان زیادی افزایش داده است (Giadinis et al. 2000). در گزارش دیگری، مصرف سلنیوم و ویتامین E پاسخ ایمنی سلولهای دام را افزایش داده است (Turner et al. 1990).
دلیل توانایی ویتامین AD3E در تحریک سیستم ایمنی با افزایش ایمونوگلوبولین مرتبط است (Al-Asadi et al., 202). Haga et al. (2021) نشان دادند که مکمل ویتامین E در گاوهای شیری، استرس اکسیداتیو را کاهش داده و پایداری غشای گلبولهای قرمز را افزایش میدهد، که ممکن است دلیل بهبودهای مشاهده شده در پارامترهای خونشناسی مستقل از تعداد پلاکتها باشد. این مقایسهها نشان میدهد که نتایج مطالعه ما با مطالعات نشخوارکنندگان تأیید شده است، و ویتامین D3 به عنوان کلید افزایش هماتوکریت و هموگلوبین ظاهر میشود.
3-4 پارامترهای تخمیر شکمبه
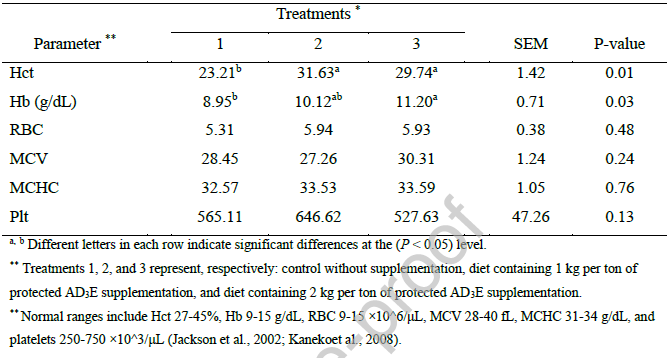
اثرات سطوح مختلف مکمل ویتامین AD3E محافظتشده بر پارامترهای تخمیر شکمبه در برههای پرواری در جدول 6 ارائه شده است. دادهها نشان میدهند که مکملسازی اثرات انتخابی و نه یکنواختی بر محیط شکمبه داشته است.
pH شکمبه در تمام تیمارها ثابت ماند و از 6.43 تا 6.67 متغیر بود (0.38 = P)، که نشان میدهد این افزودنی ظرفیت بافری شکمبه را به خطر نینداخته است. در مقابل، غلظت نیتروژن آمونیاکی (NH3-N) به طور قابل توجهی (0.01 = P) به صورت وابسته به دوز افزایش یافت. این افزایش احتمالاً نشان دهنده افزایش پروتئولیز یا فعالیت میکروبی است. در حالی که غلظت کل اسیدهای چرب فرار (TVFA) در تیمار 3 از نظر عددی در مقایسه با تیمار 2 و گروه کنترل بالاتر بود (0.05> p). قابل توجه است که نسبتهای مولی استات، پروپیونات، بوتیرات و نسبت استات به پروپیونات حاصل بدون تغییر باقی ماندند (0.05> P).
مطابق با دادههای VFA، تولید متان تخمینی نیز هیچ تفاوت معنیداری بین ... نشان نداد. این نتایج نشان میدهد که مکمل ویتامین AD3E محافظتشده در درجه اول متابولیسم نیتروژن را در شکمبه تعدیل میکند، بدون اینکه تغییر گستردهای در پروفایل اسیدهای چرب فرار یا تولید گاز ایجاد کند.
جدول 6
اثرات متفاوت مکمل ویتامین AD3E بر پارامترهای شکمبهای مشاهده شده در مطالعه حاضر - افزایش نیتروژن آمونیاکی بدون تغییرات همزمان در pH یا پروفایل اسیدهای چرب فرار - بینشهای مهمی را در مورد تعدیل انتخابی تخمیر شکمبه توسط AD3E آشکار میکند. افزایش غلظت NH3-N شکمبه با مکمل AD3E احتمالاً نشاندهنده افزایش فعالیت پروتئولیتیک در اکوسیستم میکروبی شکمبه است. این یافته با درک موجود مبنی بر اینکه آمونیاک شکمبه در درجه اول از طریق تجزیه پروتئین میکروبی، شامل باکتریهای تولیدکننده آمونیاک و فعالیت پروتوزآها تولید میشود، همسو است (Song et al., 2023a). همانطور که توسط Al-Asadi et al. (2020) پیشنهاد شده است، این کمپلکس ویتامینی ممکن است دآمیناسیون باکتریایی یا فعالیت اورهآز را تحریک کند، اگرچه مکانیسمهای خاص نیاز به بررسی بیشتر دارند. Song et al. (2023b) نشان دادند که تجویز ویتامین A در برهها، محتوای پروتئین خام در بافت ماهیچه را افزایش داده و ظرفیت آنتیاکسیدانی را بهبود میبخشد. در حالی که مطالعه آنها بر تزریق عضلانی در برههای تازه متولد شده به جای مکمل غذایی در حیوانات پرواری متمرکز بود، بهبودهای مشاهده شده در متابولیسم پروتئین نشان میدهد که ویتامین A ممکن است اثرات سیستمیکی بر مصرف نیتروژن داشته باشد. این امر از این تفسیر پشتیبانی میکند که افزایش NH3-N مشاهده شده در مطالعه حاضر میتواند نشاندهنده افزایش تجزیه پروتئین یا تغییر در توزیع نیتروژن باشد. عدم وجود تغییرات قابل توجه در pH شکمبه و پایداری نسبت اسیدهای چرب فرار اصلی، علیرغم افزایش NH3-N، نشان میدهد که مکمل AD3E متابولیسم نیتروژن را بدون ایجاد اختلال گسترده در تخمیر کربوهیدراتها تعدیل میکند.
این الگو با گزارشهایی مبنی بر تأثیر ویتامین E بر مسیرهای متابولیکی خاص - مانند سنتز اسید لینولئیک مزدوج - بدون تغییر گسترده در پروفایلهای VFA در گوسفندان مطابقت دارد (Nazari Darabkhani et al., 2024; Guerra-Rivas et al., 2016). Carvalho et al. (2024) به طور مشابه گزارش دادند که مکمل 25-هیدروکسی ویتامین D3 در گاوهای گوشتی، راندمان میکروبی شکمبه را بدون تأثیر بر pH یا نسبتهای VFA بهبود میبخشد، که نشان میدهد ویتامین D ممکن است فعالیت متابولیکی میکروبهای شکمبه را بدون تغییر پروفایلهای محصول نهایی تخمیر افزایش دهد.
این یافته با یافتههای فعلی همسو است و از این فرضیه پشتیبانی میکند که ویتامینهای AD3E میتوانند تخمیر شکمبه را تثبیت کنند و در عین حال متابولیسم نیتروژن را به طور انتخابی افزایش دهند. افزایش انتخابی NH3-N بدون تغییرات عمده در VFA پیامدهای مهمی برای درک چگونگی تأثیر مکمل ویتامین بر کارایی شکمبه دارد. غلظت آمونیاک نشان دهنده تعادل بین تجزیه پروتئین و سنتز پروتئین میکروبی است. افزایش مشاهده شده میتواند نشان دهنده افزایش پروتئولیز یا کاهش مصرف آمونیاک توسط میکروبها باشد.
مورد دوم با توجه به اینکه تولید VFA - شاخصی از فعالیت کلی تخمیر - حفظ یا افزایش یافته است، کمتر محتمل به نظر میرسد. افزایش کل VFAهای مشاهده شده در مطالعه حاضر با تحقیقات قبلی که نشان میدهد مکمل ویتامین تخمیر شکمبه را افزایش میدهد، همسو است. گزارش شده است که ویتامینهای B، C و D تجزیهپذیری ماده خشک و کارایی تخمیر را افزایش میدهند (Witariadi et al., 2022; Astawa et al., 2011).
به طور خاص، نشان داده شده است که ویتامین E به طور خطی کل اسیدهای چرب فرار (VFA)، استات و نسبت استات به پروپیونات را در شرایط آزمایشگاهی افزایش میدهد (Hou et al., 2013; Wei et al., 2015). این افزایش در تولید VFA اغلب با بهبود قابلیت هضم ماده آلی و تولید گاز همراه است (Getachew et al., 2004)، که مکانیسم قابل قبولی را برای نتایج فعلی ارائه میدهد. مکانیسمهایی که ویتامینهای محلول در چربی بر تخمیر شکمبه تأثیر میگذارند، هنوز به طور کامل مشخص نشدهاند. ویتامین A و متابولیت آن، اسید رتینوئیک، فراتر از نقشهای کلاسیک خود، دارای چندین عملکرد بیولوژیکی هستند، از جمله تعدیل بیان ژن و تمایز سلولی (Song et al., 2023a; Song et al., 2023b). در حالی که این اثرات معمولاً در بافتهای حیوانی به جای میکروبهای شکمبه مورد مطالعه قرار میگیرند، این احتمال وجود دارد که ویتامینها به طور مستقیم یا غیرمستقیم بر ترکیب جامعه میکروبی یا فعالیت متابولیکی تأثیر بگذارند.
3-5 جمعیت پروتوزوآ
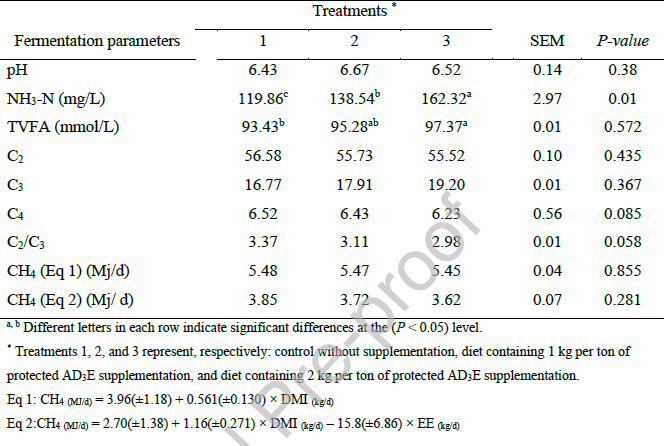
جدول 7 تأثیر سطوح مختلف مکمل ویتامین AD3E محافظتشده بر جمعیت پروتوزوآهای شکمبه برههای پرواری را نشان میدهد. پاسخ متفاوت جمعیتهای پروتوزوآ شکمبه به سطوح مختلف مکمل ویتامین AD3E محافظتشده، بهویژه اوج تعداد کل و فراوانی انتودینیوم در سطح مکمل متوسط (تیمار 2)، نشاندهنده تعامل ظریف بین ویتامینهای محلول در چربی و میکروفونای شکمبه است. غلبه انتودینیوم در بین تیمارها با نقش تثبیتشده آن به عنوان شایعترین جنس مژکدار در شکمبه سازگار است که اغلب بیش از 90٪ از کل جمعیت پروتوزوآ را تشکیل میدهد (Voia et al., 2011). مشاهده اینکه جمعیت پروتوزوآ در سطح مکمل متوسط بالاترین بود اما در بالاترین سطح کاهش یافت، نشاندهنده یک پاسخ غیرخطی و وابسته به دوز است. این الگو ممکن است اختلافات موجود در مقالات مربوط به اثرات ویتامین بر پروتوزوآ را توضیح دهد. برای مثال، مطالعهای توسط Darabkhani et al. (2024) که روشهای مختلف مکمل AD3E را در برههای پرواری بررسی میکرد، گزارش داد که تعداد پروتوزوآها به طور کلی با مکمل ویتامین کاهش یافته است. این حال، مطالعه حاضر نشان میدهد که این اثر ممکن است به دوز مصرفی بستگی داشته باشد، به طوری که سطوح متوسط به طور بالقوه محیط شکمبهای مطلوبتری را برای تکثیر پروتوزوآ ایجاد میکنند.
جدول 7
تحریک انتخابی انتودینیوم به ویژه قابل توجه است. انتودینیوم به عنوان جنس غالب، نقش مهمی در تجزیه نشاسته و عملکرد کلی شکمبه ایفا میکند (Park et al., 2018). خواص آنتیاکسیدانی ویتامین E ممکن است در این اثر نقش داشته باشد. تحقیقات نشان داده است که پروتوزوآهای بیهوازی شکمبه، از جمله انتودینیوم کاداتوم، میتوانند با موفقیت در شرایط هوازی بازدارنده، زمانی که آنتیاکسیدانهایی مانند اسید آسکوربیک و گلوتاتیون در محیط کشت گنجانده میشوند، کشت شوند (Park et al., 2018). ویتامین E (α-توکوفرول)، به عنوان یک آنتیاکسیدان قوی محلول در چربی، ممکن است به طور مشابه غشاهای پروتوزوآ ای را از آسیب اکسیداتیو در اکوسیستم شکمبه محافظت کند و در نتیجه بقا و تکثیر آنها را افزایش دهد. این اثر محافظتی با یافتههای Chaudhary et al. (2014) مطابقت دارد که گزارش دادند مکمل ویتامین E تا حدی از جمعیتهای مژکدار شکمبه در برابر اثرات سمی موننسین در بزها محافظت میکند.
مطالعات انجام شده با استفاده از کشتهای دستهای شکمبه نشان دادهاند که ویتامین E تأثیر مثبتی بر الگوی تخمیر شکمبه و جمعیت پروتوزوآ دارد (Naziroglu و همکاران، ۲۰۰۲)، اگرچه به نظر میرسد این اثرات به شدت به شکل و دوز ویتامین E مورد استفاده و همچنین نوع رژیم غذایی وابسته هستند. علاوه بر این، مکملسازی با ۵۰ واحد بینالمللی در روز ویتامین E به شکل آلفا-توکوفرول استات، با استفاده از تکنیک شبیهسازی شکمبه در شرایط آزمایشگاهی، نشان داد که به دلیل اثر آنتیاکسیدانی آن، ویتامین E جمعیت پروتوزوآ را در محیط تخمیر افزایش میدهد (Belanche et al., 2016). در مطالعه حاضر، جمعیت پروتوزوآ نیز افزایش یافت، اما سوال مهم این است که چرا این امر در شرایطی که از ویتامین E محافظتشده در شکمبه استفاده شد، رخ داد. به نظر میرسد با توجه به اینکه ویتامین مورد استفاده با روغن پالم پوشانده شده بود، در شکمبه به طور کامل محافظت نشده بود. بنابراین، اگرچه اثر آنتیاکسیدانی مثبت ویتامین E بر رشد و فعالیت پروتوزآها و در نتیجه افزایش قابلیت هضم مواد مغذی - نشان داده شده است، اما لازم است سطح محافظت ویتامینی در مطالعات آینده بهبود یابد. افزایش مشاهدهشده در قابلیت هضم پروتئین و افزایش وزن بره در کنار افزایش جمعیت پروتوزوآ با سهم شناختهشده پروتوزآها در تجزیه خوراک همسو است. پروتوزوآهای مژکدار هم بهطور مستقیم، از طریق آنزیمهای سلولولیتیک و آمیلولیتیک خود و هم بهطور غیرمستقیم، از طریق تعامل با سایر میکروبهای شکمبه، در تجزیه فیبر و نشاسته نقش دارند (Park et al., 2018). بهطور خاص، تحریک انتودینیوم ممکن است هضم نشاسته را افزایش دهد و در بهبود قابلیت هضم ذکر شده در این مطالعه نقش داشته باشد. رابطه بین پروتوزآها و تولید متان پیچیده است و صرفاً علی نیست. بهخوبی ثابت شده است که پروتوزوآها میزبان آرکیهای متانساز روی سطوح و درون سلولهای خود هستند و انتقال هیدروژن بین گونهای و متانزایی را تسهیل میکنند (Tober et al., 2024).
برخی تخمینها نشان میدهند که ۹ تا ۳۷ درصد از متانوژنها با پروتوزآها مرتبط هستند (Morgavi et al., 2012).
با این حال، عدم افزایش قابل توجه تولید متان در کنار افزایش جمعیت پروتوزآها در مطالعه حاضر کاملاً غیرمنتظره نیست. Morgavi et al. (2012) نشان دادند که رابطه بین پروتوزآها و متانزایی "یک رابطه علت و معلولی ساده" نیست، و نشان میدهند که حذف متان به طور مداوم تولید متان را کاهش نمیدهد و در برخی موارد، بسته به دوره سازگاری و تغییرات جامعه میکروبی مرتبط، میتواند آن را افزایش دهد.
وجود پروتوزآها ممکن است مسیرهای تخمیر و تقسیم هیدروژن را به روشهایی تغییر دهد که لزوماً منجر به تغییرات خطی در خروجی متان نشود. یافتههای این مطالعه هم با تحقیقات قبلی همسو و هم متفاوت است. تحریک پروتوزآها توسط مکمل AD3E با گزارشهایی مبنی بر اینکه مکملهای معدنی-ویتامینی بر جمعیت پروتوزآها تأثیری نداشتهاند (آستاوا و همکاران، ۲۰۱۱) و با کاهش کلی تعداد پروتوزآها که توسط Nazari Darabkhani et al. (2024) گزارش شده است، با مکمل AD3E در تضاد است. با این حال، نقش محافظتی ویتامین E در برابر عواملی که پروتوزآها را سرکوب میکنند توسط Chaudhary et al. (2014) پشتیبانی میشود.
علاوه بر این، توانایی آنتیاکسیدانها در حمایت از رشد پروتوزآهای بیهوازی در شرایط چالشبرانگیز، مبنایی مکانیکی برای اثر تحریکی مشاهده شده در سطوح متوسط مکمل فراهم میکند (Park et al., 2018). حداکثر پاسخ در دوز متوسط نشان میدهد که یک محدوده بهینه وجود دارد که فراتر از آن غلظتهای بالای ویتامینها ممکن است اثرات مهاری داشته باشند، احتمالاً از طریق فعالیت ضدمیکروبی مستقیم یا با تغییر ریزمحیط شکمبه به روشهایی که برای گونههای خاص پروتوزآ نامطلوب است. عدم وجود تغییرات قابل توجه در سایر جنسها (Ophryoscolex، Diplodinium، Isotricha، Dasytricha) نشان میدهد که Entodinium ممکن است به طور خاص به شرایط ایجاد شده توسط مکمل AD3E واکنش نشان دهد، که ماهیت خاص این تعامل را برجسته میکند.
3-6 ویژگیهای لاشه
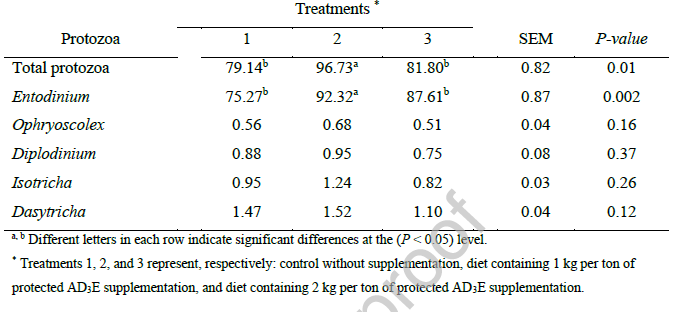
ارزیابی ویژگیهای لاشه در برههای پرواری که با ویتامین AD3E محافظتشده مکملدهی شده بودند، هیچ اثر درمانی معنیداری بر هیچ یک از پارامترهای اندازهگیری شده نشان نداد (جدول 8). وزن زنده در زمان کشتار، وزن لاشه، اجزای غیر لاشهای (پشم و پوست) و اندازهگیریهای بیومتریک (طول، عرض، طول ران، طول دست) از نظر آماری در سطوح مکملدهی بدون تغییر باقی ماندند. این نتایج نشان میدهد که بهبود عملکرد زنده مشاهده شده در این مطالعه (جدول 2) به تغییرات قابل تشخیص در عملکرد لاشه یا صفات ساختاری منجر نشده است. دلیل اینکه وزن کشتار (جدول 8) برخلاف وزن نهایی حیوانات (جدول 2) معنیدار نیست، این است که ما سه حیوان بزرگ را از هر گروه انتخاب کردیم و دادههای وزن قبل از کشتار با دادههای وزن نهایی در جدول 2 متفاوت است.
جدول 8
عدم وجود اثرات لاشه با مجموعه قابل توجهی از مقالات علمی که مکمل ویتامین در نشخوارکنندگان را بررسی میکنند، همسو است. Andrade et al. (2023) ترکیبات ویتامین محلول در آب و محلول در چربی (از جمله ADE) را در گاوهای نر جوان نلور در یک دوره ۱۷۰ روزه ارزیابی کردند و هیچ اثر معنیداری بر صفات لاشه گزارش نکردند و نتیجه گرفتند که مکمل ویتامین بر ویژگیهای لاشه در گاوهای پرواری تأثیر نمیگذارد. این یافته مستقیماً از مشاهدات مطالعه حاضر پشتیبانی میکند و تعمیمپذیری این پدیده را در گونههای نشخوارکنندگان و سیستمهای تولیدی گسترش میدهد.
به طور مشابه، تحقیقاتی که به طور خاص مکمل ویتامین E را در برهها بررسی کردهاند، به طور مداوم اثرات حداقلی بر صفات لاشه گزارش کردهاند. Atay et al. (2009) مکمل ویتامین E را در برههای نر کاریا بررسی کردند و علیرغم برخی بهبودها در پایداری گوشت و افزایش عددی در راندمان تبدیل خوراک، هیچ اثر معنیداری بر ویژگیهای لاشه نیافتند.
مطالعات انجام شده روی برههای آواسی این یافتهها را تأیید کرده و نشان میدهد که اگرچه ویتامین E ممکن است بر ویژگیهای کیفی گوشت مانند اکسیداسیون لیپید، کاهش چربی و پایداری رنگ در طول ذخیرهسازی تأثیر مثبت داشته باشد، اما وزن یا ابعاد لاشه را به طور قابل توجهی تغییر نمیدهد (Macit et al., 2003).
این تفکیک بین بهبود عملکرد و عدم وجود اثرات لاشه با بخش قابل توجهی از ادبیات بررسی مکمل ویتامین در نشخوارکنندگان همسو است. Andrade et al. (2023) ترکیبات ویتامین محلول در آب و محلول در چربی (از جمله ADE) را در گاوهای نر جوان نلور در یک دوره ۱۷۰ روزه ارزیابی کردند و هیچ تأثیر قابل توجهی بر صفات لاشه گزارش نکردند و نتیجه گرفتند که مکمل ویتامین بر ویژگیهای لاشه در گاوهای پرواری تأثیر نمیگذارد. این یافته مستقیماً از مشاهدات مطالعه حاضر پشتیبانی میکند و تعمیمپذیری این پدیده را در گونههای نشخوارکنندگان و سیستمهای تولیدی گسترش میدهد. به طور مشابه، تحقیقاتی که به طور خاص مکمل ویتامین E را در برهها بررسی کردهاند، همواره اثرات حداقلی بر ویژگیهای لاشه گزارش کردهاند.
Atay et al. (2009) مکمل ویتامین E را در برههای نر کاریا بررسی کردند و علیرغم برخی بهبودها در پایداری گوشت و افزایش عددی در راندمان تبدیل خوراک، هیچ اثر معنیداری بر ویژگیهای لاشه نیافتند. مطالعات انجام شده بر روی برههای آواسی این یافتهها را تأیید کرده و نشان میدهد که اگرچه ویتامین E ممکن است بر ویژگیهای کیفی گوشت مانند اکسیداسیون لیپید، از دست دادن آب و پایداری رنگ در طول ذخیرهسازی تأثیر مثبت داشته باشد، اما وزن یا ابعاد لاشه را به طور قابل توجهی تغییر نمیدهد (Macit et al., 2003). این تفکیک بین افزایش عملکرد و اصلاح لاشه نشان میدهد که ویتامینها ممکن است اثرات اولیه خود را بر راندمان متابولیک و کیفیت بافت اعمال کنند تا مورفولوژی لاشه.
یافتههای حاضر همچنین با تحقیقات در مورد فناوریهای مواد مغذی محافظتشده سازگار است.
Behan et al. (2019)، در یک بررسی جامع از مکملهای چربی محافظتشده در شکمبه در حیوانات شیری، خاطرنشان کردند که اگرچه مواد مغذی محافظتشده باعث افزایش مصرف انرژی و بهبود وضعیت بدن میشوند، اما به طور مداوم ویژگیهای لاشه را تغییر نمیدهند. این تشابه بین چربیهای محافظتشده و ویتامینهای محافظتشده نشان میدهد که سیستمهای تحویل محافظتشده در شکمبه ممکن است کارایی متابولیکی و عملکرد تولیدی را بدون نیاز به هدایت مجدد مواد مغذی به سمت رسوب لاشه، در اولویت قرار دهند. عدم پاسخ لاشه در مطالعه حاضر با شواهد نوظهور مبنی بر اینکه متابولیتهای ویتامین D میتوانند بر ویژگیهای لاشه در گاو تأثیر بگذارند، در تضاد است.
Carvalho et al. (2024) نشان دادند که مکمل ۲۵-هیدروکسی ویتامین D3 در گاوهای نر نلور، درصد پوشش لاشه را افزایش داده (۵۵.۲٪ در مقابل ۵۳.۸٪) و تمایل به افزایش سطح عضله Longissimus دارد، بدون اینکه بر مرمری شدن یا درجه عملکرد تأثیر بگذارد. این اثر به افزایش بیان ژنهای مرتبط با رشد عضلات اسکلتی، از جمله فاکتور رشد شبه انسولین-۱ (IGF1)، IGF2 و هدف پستانداران راپامایسین (mTOR؛ Martins et al., 2025) نسبت داده شد. اختلاف بین این یافتهها و مطالعه حاضر ممکن است نشاندهنده تفاوتهای گونهای (گاو در مقابل گوسفند)، متابولیت خاص ویتامین D مورد استفاده (۲۵-هیدروکسی ویتامین D3 در مقابل ویتامین D3 استاندارد) یا ملاحظات دوز باشد.
مودادو و همکاران (۲۰۲۴) همچنین گزارش دادند که مکمل ۲۵-هیدروکسی ویتامین D3 در گاوهای نر جوان نلور منجر به افزایش ۸ کیلوگرمی وزن حیوانات نسبت به گروه کنترل بدون مکمل و افزایش بیشتر در عرض دندهها شد که نشاندهنده رشد بیشتر لاشه است.
این یافتهها نشان میدهد که شکل و سطح مکمل ویتامین و همچنین مرحله فیزیولوژیکی حیوان ممکن است از عوامل تعیینکننده مهم پاسخ لاشه باشند. در نشخوارکنندگان کوچک، شواهد محدودی نشاندهنده پتانسیل بهبود لاشه در شرایط خاص است. Mudado et al. (2024) گزارش دادند که مکمل ویتامین ترکیبی در بزها، ویژگیهای لاشه، از جمله افزایش ۸ تا ۱۰ درصدی توده عضلانی را بهبود بخشید، که نشان میدهد سطوح بالاتر مکمل یا ترکیبات خاص ممکن است، بر نتایج لاشه در نشخوارکنندگان کوچک تأثیر بگذارد. عدم وجود چنین اثراتی در مطالعه حاضر، مفهوم یک بازه دوز بهینه را تقویت میکند، که در آن مکمل متوسط ممکن است عملکرد زنده را افزایش دهد در حالی که سطوح بالاتر برای تبدیل افزایش رشد به مزایای قابل اندازهگیری لاشه مورد نیاز است.
3-7 هیستومورفومتری روده
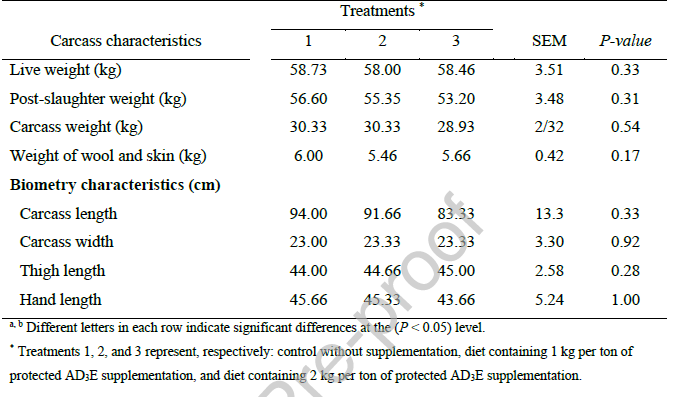
ارزیابی هیستومورفولوژی روده کوچک (جدول 9) هیچ اثر درمانی معنیداری بر طول پاپیلا، عرض پاپیلا یا عمق کریپت در دوازدهه، ژژنوم و ایلئوم نشان نداد (P > 0.05). این یافتهها نشان میدهد که مکمل ویتامین AD3E محافظتشده، در سطوح مورد آزمایش، ساختار ماکروسکوپی روده کوچک را در برههای پرواری به طور قابل توجهی تغییر نمیدهد. عدم تغییر مورفولوژیکی ممکن است نشان دهد که کفایت تغذیهای پایه برای حفظ ساختار روده کافی بوده است، یا اینکه AD3E بر پارامترهای عملکردی - مانند تحرک، جذب مواد مغذی یا تعاملات میکروبیوم - تأثیر گذاشته است، نه بر 3-ویژگیهای ساختاری ناخالص. تحقیقات نشان داده است که ویتامینهای A، D3 و E میتوانند تحرک روده را تعدیل کرده و بر جمعیتهای میکروبی روده تأثیر بگذارند، که برای حفظ سلامت و عملکرد روده ضروری هستند (Malaguarnera، 2020). عدم وجود تغییرات مورفولوژیکی قابل توجه در روده کوچک پس از مصرف مکمل ویتامین AD3E محافظتشده، با شواهد رو به رشدی مطابقت دارد که نشان میدهد اثرات اولیه ویتامینهای محلول در چربی بر دستگاه گوارش، به ویژه در حیوانات سالم و با تغذیه کافی، ممکن است عملکردی و تعدیلکننده سیستم ایمنی باشد تا ساختاری.
جدول 9
یافتههای حاضر با تحقیقاتی که نشان میدهند ویتامین D نقش نقشی در هموستازه و تنظیم میکروبیوتا بدون تغییر لزوماً در معماری روده ایفا میکند، مطابقت دارد.Malaguerra (2020) گفت که مسیرهای ویتامین D به طور حیاتی در هموستاز روده دخیل هستند و این ویتامین به طور قابل توجهی برای ترکیب میکروبیوتای روده و فعالیتهای پویای آن تاثیر میگذارد. این نقش تعدیلکننده سیستم ایمنی ممکن است از تغییرات در ارتفاع پرزها یا عمق کریپتها رخ دهد، که از این تفسیر پشتیبانی میکند که عدم وجود تغییر مورفولوژیکی در مطالعه حاضر، عملکرد عملکردی را از بین نمیبرد. علاوه بر این، Wu et al. (2022b) به طور جامع رابطه بین میکروبیوتای روده و عملکرد کلسیم را بررسی کردند و بررسی کردند که ویتامین D با فلور روده و عوامل مرتبط با اسیدهای چرب زنجیره کوتاه (SCFAs) و عواملی برای تأثیرگذاری بر فیزیولوژیک میزبان عناصر دارد. این عوامل میتوانند جذب مواد مغذی و عملکرد سدی را بدون نیاز به تغییرات قابل اندازهگیری در ابعاد مختلف، افزایش دهند.
در حالی که مطالعه حاضر هیچ تغییر مورفولوژیکی با مکمل AD3E پیدا نکرد، شواهد حاصل از سایر مدلهای تجربی نشان میدهد که ویتامین E میتواند در شرایط چالش فیزیولوژیکی، ساختار روده را تحت تأثیر قرار دهد. Shirpoor et al. (2006) تأثیر ویتامین E را بر تغییرات رودهای ناشی از دیابت در موشها بررسی کردند و دریافتند که حیوانات دیابتی افزایش قابل توجهی در ارتفاع پرزها، عمق کریپتها و ضخامت لایه عضلانی نشان دادند - تغییراتی که با مکمل ویتامین E کاملاً به حالت عادی برگشتند. نویسندگان این اثر محافظتی را به افزایش ظرفیت آنتیاکسیدانی پلاسما و کاهش پراکسیداسیون لیپید نسبت دادند، که نشان میدهد ویتامین E بیشترین اثرات ساختاری خود را زمانی که روده تحت استرس اکسیداتیو قرار دارد، اعمال میکند. این یافته به ویژه برای تفسیر نتایج صفر فعلی مرتبط است: در برههای سالم و بدون استرس با تغذیه پایه کافی، ظرفیت آنتیاکسیدانی ویتامین E ممکن است برای حفظ هموستاز بدون ایجاد سازگاریهای ساختاری قابل اندازهگیری کافی باشد. نقش محافظتی ویتامین E در شرایط چالش برانگیز بیشتر توسط Vijayalakshmi et al. (2005) پشتیبانی میشود، که نشان داد محدودیت ویتامین در موشها به طور قابل توجهی آپوپتوز سلولهای اپیتلیال روده را افزایش داده و معماری پرزها را به خطر میاندازد، در حالی که مکمل ویتامین E این تغییرات را معکوس میکند. نویسندگان نتیجه گرفتند که "به نظر میرسد افزایش استرس اکسیداتیو بافت، آپوپتوز ناشی از محدودیت ویتامین در سلولهای اپیتلیال روده در موشها را واسطهگری میکند" و ویتامین E، از طریق خواص آنتیاکسیدانی خود، به ویژه در بازگرداندن یکپارچگی روده مؤثر است.
تحقیقات آینده باید شامل اندازهگیری نفوذپذیری روده، نشانگرهای ایمنی مخاطی، ترکیب جامعه میکروبی و بیان ژن ناقل مواد مغذی باشد تا پیامدهای عملکردی مکمل AD3E را به طور کامل مشخص کند. علاوه بر این، بررسی اثرات این ویتامینها در شرایط استرس فیزیولوژیکی یا تغذیه نامطلوب ممکن است مزایای ساختاری را آشکار کند که در حیوانات سالم آشکار نیست.
- نتیجهگیری
استفاده از ویتامینهای محافظتشده AD3E در سطح متوسط (۱ کیلوگرم در هر تن) به طور قابل توجهی عملکرد رشد و قابلیت هضم پروتئین را در برههای پرواری بهبود میبخشد. این ویتامین از طریق افزایش متابولیسم نیتروژن در شکمبه، افزایش غلظت نیتروژن آمونیاکی و تحریک انتخابی جمعیتهای مفید پروتوزوآیی، به ویژه انتودینیوم، بدون ایجاد اختلال در pH شکمبه، پروفایل اسیدهای چرب فرار یا تولید متان، عملکرد رشد را ۲.۵ کیلوگرم دیگر افزایش میدهد.
ویتامین AD3E همچنین پارامترهای خونشناسی را بهبود میبخشد که نشاندهنده کاهش استرس اکسیداتیو و افزایش پایداری گلبولهای قرمز در برههای دریافتکننده ویتامین است. با این حال، این بهبودهای متابولیکی و عملکردی منجر به تغییر در ویژگیهای لاشه یا مورفولوژی روده نمیشود. یافتههای این تحقیق نشان میدهد که افزودن AD3E به میزان ۱ تا ۲ کیلوگرم در هر تن، عملکرد شکمبه و سلامت سیستمیک برههای پرواری را بهبود میبخشد و در نهایت رشد بره را ۲.۵ کیلوگرم افزایش میدهد، بدون اینکه تغییرات ساختاری در لاشه یا بافتهای روده ایجاد کند.
منابع تأمین مالی
این تحقیق هیچ کمک مالی خاصی از سازمانهای تأمین مالی در بخشهای دولتی، تجاری یا غیرانتفاعی دریافت نکرده است.
سپاسگزاری
ما صمیمانه از شرکت بهدام رشد خراسان (ایران) برای ارائه سخاوتمندانه مکملهای ویتامینی مورد نیاز برای این مطالعه تحقیقاتی قدردانی میکنیم. حمایت آنها در امکان پذیر کردن این تحقیق علمی نقش مهمی داشت.
جدول 1: اجزا و ترکیبات شیمیایی جیرههای غذایی مورد استفاده در آزمایش
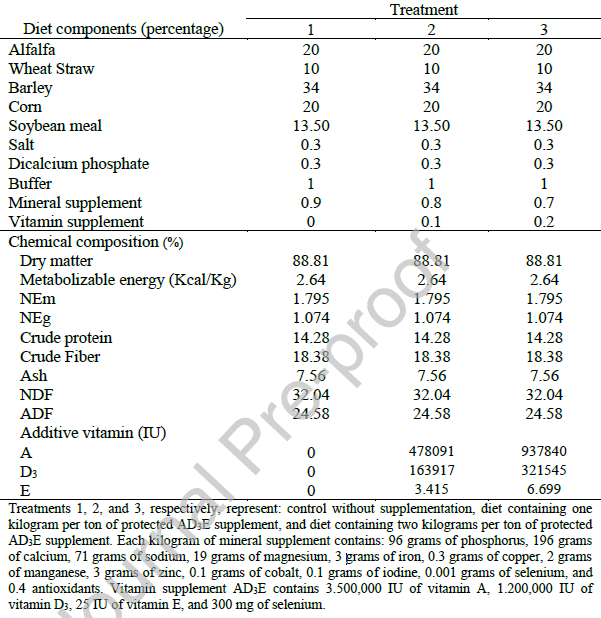
جدول ۲: تأثیر سطوح مختلف مکمل ویتامین AD3E محافظتشده بر عملکرد برههای پرواری.
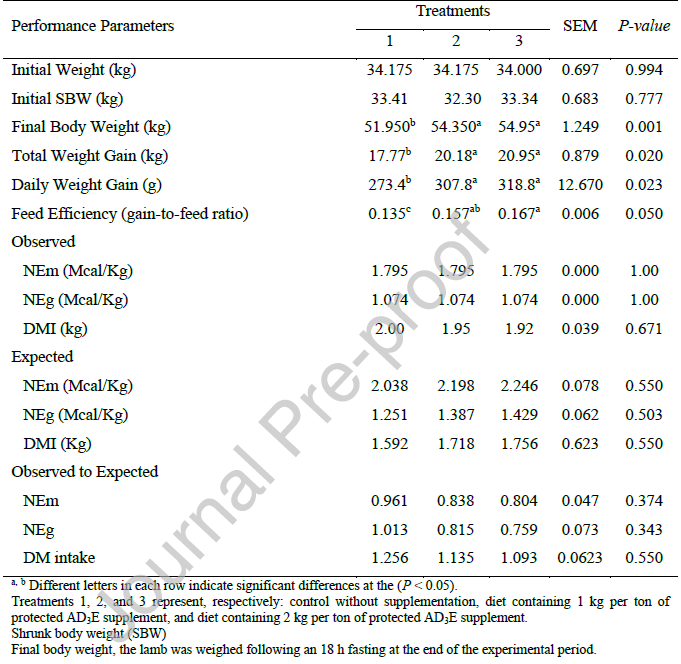
جدول 3: تأثیر سطوح مختلف مکمل ویتامین AD3E محافظتشده بر قابلیت هضم خوراک.
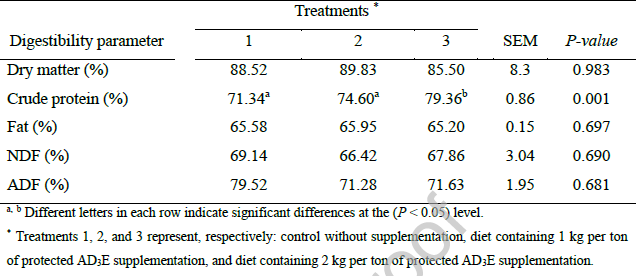
جدول 4: تأثیر سطوح مختلف مکمل ویتامین AD3E محافظتشده بر پارامترهای بیوشیمیایی خون و آنزیمهای کبدی برههای پرواری.
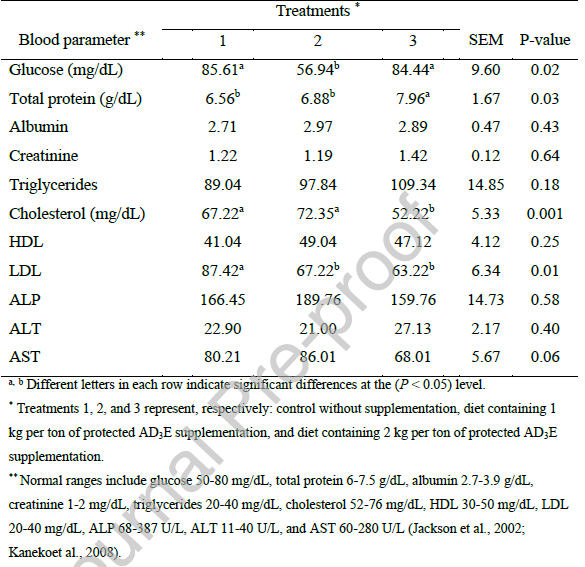
جدول ۵. تأثیر سطوح مختلف مکمل ویتامین AD3E محافظتشده بر هماتولوژی برههای پرواری.
جدول 6: تأثیر سطوح مختلف مکمل ویتامین AD3E محافظتشده بر پارامترهای تخمیر شکمبه.
جدول 7: تأثیر سطوح مختلف مکمل ویتامین AD3E محافظتشده بر جمعیت پروتوزوآ (N×105) در شکمبه برههای پرواری.
جدول ۸: تأثیر سطوح مختلف مکمل ویتامین AD3E محافظتشده بر ویژگیهای لاشه برههای پرواری.
جدول ۹: تأثیر سطوح مختلف مکمل ویتامین AD3E محافظتشده بر مورفولوژی روده برههای پرواری.
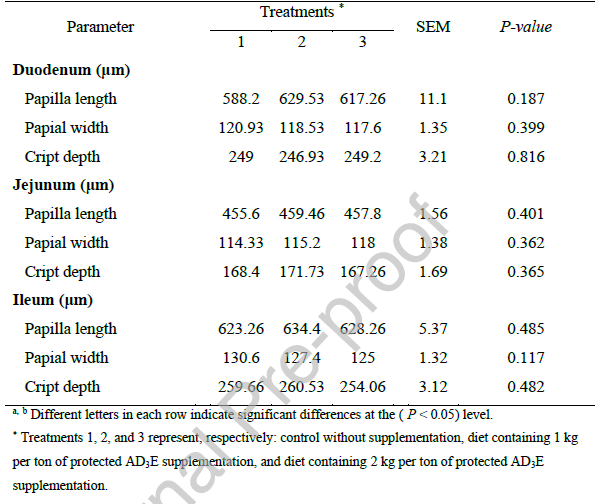
شکل 1- تأثیر سطوح مختلف مکمل ویتامین AD3E محافظتشده بر میزان مالوندیآلدئید خون در برههای پرواری.
شکل ۲. تأثیر سطوح مختلف مکمل ویتامین AD3E محافظتشده بر تعداد لکوسیتها و نسبت نوتروفیل به لنفوسیت در برههای پرواری.
منابع
Al-Asadi, F. A. A., Habib, H. N., & Hassan, A. F. (2020). Effect of the injection of vitamins AD3E and the seasons on some blood traits, biochemical components, and hormones of Arabi rams. Journal of Animal Behaviour and Biometeorology, 8(1), 74-81. https://doi.org/10.31893/jabb.20010
Al_Galiby, M. K. A., & Al-khafaji, H. Y. S. (2022). Studying the effect of giving doses of different levels of the group of vitamins AD3E on some productive and doll traits For young male goats the local. University of Thi-Qar Journal of agricultural research, 11(2), 187-196. https://doi.org/10.54174/utjagr.v11i2.198
Amevor, F. K., Cui, Z., Du, X., Ning, Z., Deng, X., Xu, D., Shu, G., Wu, Y., Cao, X., Shuo, W., Tian, Y., Li, D., Wang, Y., Zhang, Y., Du, X., Zhu, Q., Han, X. & Zhao, X. (2022). Supplementation of dietary quercetin and vitamin E promotes the intestinal structure and immune barrier integrity in aged breeder hens. Frontiers in Immunology, 13, 860889. https://doi.org/10.3389/fimmu.2022.860889Andrade, D. R., Silva, F. A. S., Pinheiro, J. S., da Silva, J. T., Trópia, N. V., Godoi, L. A., Vilela, R. S. R., Cidrini, F. A. A., Rennó, L. N., Zanetti, D., Acedo, T. S., & Valadares Filho, S. C. (2023). Does vitamin blends supplementation affect the animal performance, carcass traits, and nutrient digestibility of young Nellore finishing bulls? Animal Bioscience, 36(12), 1831- 1841. https://doi.org/10.5713/ab.23.0087
Asadian, A., Mezes, M., & Mirhadi, S. A. (1996). Effect of vitamins A and E on blood plasma vitamin status and daily body mass gain of different fat-tailed sheep breeds. Acta Veterinaria Hungarica, 44(1), 99-109. https://pubmed.ncbi.nlm.nih.gov/8826705
Astawa, P. A., Partama, I. B. G., Suyadnya, P., & Sutarpa, I. N. S. (2011). Effect of vitaminmineral supplementation in commercial feed on the digestibility coefficient and rumen fermentation of Bali cattle. Journal of the Indonesian Tropical Animal Agriculture, 36(1), 69- 74. https://doi.org/10.14710/jitaa.36.1.69-74
Atay, O. K. A. N., Gökdal, Ö. Z. D. A. L., Eren, V. A. D. U. L. L. A. H., Çetiner, Ş. E. V. K. İ., & Yikilmaz, H. (2009). Effects of dietary vitamin E supplementation on fattening performance, carcass characteristics and meat quality traits of Karya male lambs. Archives Animal Breeding, 52(6), 618-626. https://doi.org/10.5194/aab-52-618-2009
Baldin, S.R., Millen, D.D., Martins, C.L., Pereira, A.S.C., Barducci, R.S., & de Beni, M.A.(2013). Desempenho, características de carcaça e carne de bovinos nelore e canchim, confinados com dietas suplementadas com vitaminas D e E. Acta Science - Animal Science. 35, 403–410. https://doi.org/10.4025/actascianimsci.v35i4.18801.
Belanche, A., Kingston-Smith, A. H., & Newbold, C. J. (2016). An integrated multi-omics approach reveals the effects of supplementing grass or grass hay with vitamin E on the rumen microbiome and its function. Frontiers in microbiology, 7, 905.
Behan, A. A., Loh, T. C., Fakurazi, S., Kaka, U., Kaka, A., & Samsudin, A. A. (2019). Effects of
supplementation of rumen protected fats on rumen ecology and digestibility of nutrients in
sheep. Animals, 9(7), 400. https://doi.org/10.3390/ani9070400
Broderick, G.A., & Kang, J.H. (1980). Automated simultaneous determination of ammonia and
total amino acids in ruminal fluid and in vitro media. Journal of Dairy Science, 63, 64-75.
https://doi.org/10.3168/jds.S0022-0302(80)82888-8
Cantorna, M. T., Snyder, L., & Arora, J. (2019). Vitamin A and vitamin D regulate the microbial
complexity, barrier function, and the mucosal immune responses to ensure intestinal
homeostasis. Critical Reviews in Biochemistry and Molecular Biology. 54(2):184-192.
https://doi.org/10.1080/10409238.2019.1611734
Cao, G., Yu, Y., Wang, H., Yang, H., Tao, F., Yang, S., Liu, J., Li, Z., & Yang, C. (2024).
Dietary Clostridium butyricum and 25-Hydroxyvitamin D3 modulate bone metabolism of
broilers through the gut-brain axis. Poultry Science, 103(8):103966. https://doi.org/10.1016/j.psj.2024.103966
Carvalho, P. H. V., Oh, J., Estill, C. T., Wiegand, J. B., Cooke, R. F., Cappellozza, B. I., Pohler,
K. G., Starkey, J. D., Hall, J. B., & Starkey, J. D. (2024). The interaction of feeding an eubiotic blend of essential oils plus 25-hydroxy-vitamin D3 on performance, carcass characteristics, and immune response of finishing beef cattle compared with monensin and supplemental vitamin E. Journal of Animal Science, 102, skad414. https://doi.org/10.1093/jas/skad414
Chaudhary, U. B., Upadhyaya, R. S., & Nawab Singh, N.S. (2014). Effect of feeding monensin
and vitamin E on rumen ciliate population in goats. The Indian Journal of Animal Sciences,
84(5), 527-530. https://epubs.icar.org.in/index.php/IJAnS/article/view/39256
Chen, G. (2021). The interactions of insulin and vitamin A signaling systems for the regulation
of hepatic glucose and lipid metabolism. Cells, 10(8), 2160. https://doi.org/10.3390/cells10082160
Cheung, L. Y., & Camper, S. A. (2020). PROP1-dependent retinoic acid signaling regulates
developmental pituitary morphogenesis and hormone expression. Endocrinology, 161(2), bqaa002. https://doi.org/10.1210/endocr/bqaa002
Dakshinamurti, K. (2015). Vitamins and their derivatives in the prevention and treatment of metabolic syndrome diseases (diabetes). Canadian Journal of Physiology and Pharmacology, 93(5), 355–362. https://doi.org/10.1139/cjpp-2014-0479
de Andrade, D. R., de Sales Silva, F. A., de Souza Pinheiro, J., da Silva, J. T., Trópia, N. V., Godoi, L. A., ... & de Campos Valadares Filho, S. (2023). Does vitamin blends supplementation affect the animal performance, carcass traits, and nutrient digestibility of young Nellore finishing bulls?. Animal Bioscience, 36(12), 1831. https://doi.org/10.5713/ab.23.0087
Dehority, B. A. (2003). Rumen Microbiology. Nottingham University Press, Nottingham, UK. El-Sabban, F. F., Rothenbacher, H., Long, T. A., & Baumgardt, B. R. (1971). Certain blood constituents and serum transaminases in Hereford steers fed high-energy rations. American Journal of Veterinary Research, 32 (7), 1027-1032. https://doi.org/10.2460/ajvr.1971.32.07.1027
Ferdouse, S., Hossain, M. S., Talukder, R., Lovelu, M. A., Rahman, N., & Nath, S. K. (2024). Effects of dietary supplementation with vitamin A, D3, E, and garlic on heat detection, pregnancy rate, and serum profile in anestrus dairy cows. Veterinary and Animal Science, 26, 100396. https://doi.org/10.1016/j.vas.2024.100396
Fu, Z., Noguchi, T. & Kato, H. (2001). Vitamin A deficiency reduces insulin-like growth factor (IGF)-I gene expression and increases IGF-I receptor and insulin receptor gene expression in tissues of Japanese quail (Coturnix coturnix japonica). Journal of Nutrition, 131 (4), 1189– 1194. https://doi.org/10.1093/jn/131.4.1189
Getachew, G., DePeters, E. J., & Robinson, P. H. (2004). In vitro gas production provides effective method for assessing ruminant feeds. California Agriculture, 58(1), 54-58. https://doi.org/10.3733/ca.v058n01p54
Giadinis, N., Koptopoulos, G., Roubies, N., Siarkou, V., & Papasteriades, A. (2000). Selenium and vitamin E effect on antibody production of sheep vaccinated against enzootic abortion (Chlamydia psittaci). Comparative Immunology, Microbiology and Infectious Diseases, 23(2), 129-137. https://doi.org/10.1016/S0147-9571(99)00066-1
Gorocica-Buenfil, M.A., Fluharty, F.L., Bohn, T., Schwartz, S.J., & Loerch, S.C.(2007). Effect of low vitamin A diets with high-moisture or dry corn on marbling and adipose tissue fatty acid composition of beef steers. Journal Animal Science. 85, 3355–3366. https://doi.org/10.2527/1997.753604x
Guerra-Rivas, C., Gallardo, B., Mantecón, Á. R., del Álamo, A. G., Lavín, P., & Manso, T. (2017). Evaluation of protective effect of different dietary fiber on polyphenolic profile bioavailability in rats. Journal of the Science of Food and Agriculture, 97(8), 2531-2538. https://doi.org/10.1002/jsfa.8070
Guerra-Rivas, C., Vieira, C., Rubio, B., Martínez, B., Gallardo, B., Mantecón, A. R., ... & Manso, T. (2016). Effects of grape pomace in growing lamb diets compared with vitamin E and grape seed extract on meat shelf life. Meat science, 116, 221-229. https://doi.org/10.1016/j.meatsci.2016.02.022
Luan, J., Jin, Y., Zhang, T., Feng, X., Geng, K., Zhang, M., & Geng, C. (2023). Effects of dietary vitamin E supplementation on growth performance, slaughter performance, antioxidant capacity and meat quality characteristics of finishing bulls. Meat Science, 206, 109322. https://doi.org/10.1016/j.meatsci.2023.109322
Habeeb, A. A. M., AbdelHafez, M. A. M., ELGohary, E. S. H., Fathala, M. M., & Salama, O. A. (2015). Effect of vitamin AD3E injection on age and weight of weaning and reproductive activity of goats. 1- physiological response and reproductive performance of goat bucks during different seasons in Egypt. Journal of Animal and Poultry Production, 6(12), 719-739. https://doi.org/10.21608/jappmu.2015.52956
Habeeb, A. A. M., El-Gohary, E. S., Saleh, H. M., & El-Deep, M. M. (2008). Effect of summer heat stress conditions and feeding protein level on milk yield and composition in Ossimi ewes and their lambs performance. Egyptian Journal of Applied Sciences, 23, 409-429.
Habeeb, A. A., El-Tarabany, A. A., Gad, A. E., & Atta, M. A. (2018). Negative effects of heat stress on physiological and immunity responses of farm animals. Change, 16(2.0), 0-2. https://doi.org/10.31058/j.as.2018.21001
Haga, S., Ishizaki, H., & Roh, S. (2021). The physiological roles of vitamin E and hypovitaminosis E in the transition period of high-yielding dairy cows. Animals, 11(4), 1088. https://doi.org/10.3390/ani11041088
Hernández, J. M. & Tamassia, L. (2025). Optimizing ruminant health through optimum vitamin nutrition. Animal Nutrition and Health. Retrieved from https://www.dsmfirmenich. com/anh/news/feed-talks/articles/optimizing-ruminant-health-through-ovn.html. Assessed in Feb 13, 2026.
Keywanloo, M., Ahmadi-Hamedani, M., Jebelli Javan, A., & Rakhshani Zabol, F. (2021). Effects of parenteral vitamin D3 supplementation on Hematological Parameters of Healthy Holstein Bulls. Archives of Razi Institute, 76(5), 1509. https://doi.org/10.22092/ari.2020.352209.1552
Hou, J., Wang, F., Wang, Y., & Liu, F. (2013). Effects of vitamin E on the concentration of conjugated linoleic acids and accumulation of intermediates of ruminal biohydrogenation in vitro. Small Ruminant Research, 111(1-3), 63-70. https://doi.org/10.1016/j.smallrumres.2012.09.015
Ismail Mahmud, K., & Karadas, F. (2023). Effects of the melatonin and vitamin e (alphatocopherol acetate) as antioxidants on biochemical blood parameters, lipid profile, and muscle vitamin e concentration in Awassi lambs fed a high-energy diet and normal diet. Archives of Razi Institute, 78(1), 249–259. https://doi.org/10.22092/ARI.2022.358494.2234
Jackson, P. G., Cockcroft, P. D., & Elmhurst, S. (2002). Clinical examination of farm animals (Vol. 331). Oxford: Blackwell Science.
Kaneko, J.J., Harvey, J.W. and Bruss, M. (2008) Clinical Biochemistry of Domestic Animals. 6th Edition, Elsevier Inc., Academic Press, Cambridge, 117-138.
Kannan, G., Terrill, T. H., Kouakou, B., & Lee, J. H. (2024). Influences of a supplemental blend of essential oils plus 25-hydroxy-vitamin D3 on growth performance of meat goats. Animals, 14(9), 1391. https://doi.org/10.3390/ani14091391
Khan, M. Z., Huang, B., Kou, X., Chen, Y., Liang, H., Ullah, Q., Khan, I. M., Khan, A., Chai, W., & Wang, C. (2024). Enhancing bovine immune, antioxidant and anti-inflammatory responses with vitamins, rumen-protected amino acids, and trace minerals to prevent periparturient mastitis. Frontiers in Immunology, 14, 1290044. https://doi.org/10.3389/fimmu.2023.1290044
Kiani, A., & Nielsen, M. O. (2022). Metabolic and hematological responses to fasting and transportation in lambs. Journal of Animal Physiology and Animal Nutrition, 106(3), 567- 578. https://doi.org/10.1111/jpn.13645
Kitchenham, B. A., Rowlands, G. J., Manston, R., & Dew, S. M. (1975). The blood composition of dairy calves reared under conventional and rapid-growth systems. British Veterinary Journal, 131(4), 436-446. https://doi.org/10.1016/s0007-1935(17)35239-9
Klasing, K. C. (2007). Nutrition and the immune system. British Poultry Science, 48(5), 525- 537. https://doi.org/10.1080/00071660701671336
Knutson, E. E., Menezes, A. C. B., Sun, X., Fontoura, A. B. P., Liu, J. H., Bauer, M. L., ... & Ward, A. K. (2020). Effect of feeding a low-vitamin A diet on carcass and production characteristics of steers with a high or low propensity for marbling. Animal, 14(11), 2308- 2314. https://doi.org/10.1017/S1751731120001135
Li, Y. J., Li, L. Y., Li, J. L., Zhang, L., Gao, F., & Zhou, G. H. (2015). Effects of dietary supplementation with ferulic acid or vitamin e individually or in combination on meat quality and antioxidant capacity of finishing pigs. Asian-Australasian Journal of Animal Sciences, 28(3):374-381. https://doi.org/10.5713/ajas.14.0432
Lobo-Jr, A.R., Delgado, E.F., Mourão,G.B., Pedreira, A.C.M.S., Berndt,B., & Demarchi, J.J.A.A. (2012).Interaction of dietary vitamin D3 and sunlight exposure on B. indicus cattle:.Animal performance, carcass traits, and meat quality. Livestock Science. 145:96–204. doi:10.1016/j.livsci.2012.02.002
Malaguarnera, L. (2020). Vitamin D and microbiota: Two sides of the same coin in the immunomodulatory aspects. International Immunopharmacology, 79, 106112. https://doi.org/10.1016/j.intimp.2019.106112
Martins, T. E., Gouvêa, V. N., Perdigão, A., Niehues, M. B., Martins, C. L., Millen, D. D., Acedo, T. S., Carvalho, V. V., Tamassia, L. F. M., & Arrigoni, M. D. B. (2025). Effects of supplemental 25-hydroxyvitamin D3 on growth performance, physiological responses, and gene expression of skeletal muscle growth of finishing beef cattle. Journal of Animal Science, 103, skaf090. https://doi.org/10.1093/jas/skaf090
McCullough, F. S. W., Northrop-Clewes, C. A., & Thurnham, D. I. (1999). The effect of vitamin A on epithelial integrity. Proceedings of the Nutrition Society, 58(2), 289-293. https://doi.org/10.1017/s0029665199000403
McDonald, P., Edwards, R. A., Greenhalgh, J. F. D., Morgan, C. A., Sinclair, L. A., & Wilkinson, R. G. (2010). Animal Nutrition (7th ed.). Prentice Hall/Pearson.
McDowell, M. A., Fryar, C. D., Ogden, C. L., & Flegal, K. M. (2008). Anthropometric reference data for children and adults: United States, 2003–2006. National health statistics reports, 10(1-45), 5. https://www.cdc.gov/nchs/data/nhsr/nhsr010.pdf
Morgavi, D. P., Martin, C., Jouany, J. P., & Ranilla, M. J. (2012). Rumen protozoa and methanogenesis: Not a simple cause–effect relationship. British Journal of Nutrition, 107(3), 388-397. https://doi.org/10.1017/S0007114511002935
Mudado, F. S., Silveira, M. B., Fernandes, H. J., Perdigão, A., dos Santos, D. M. M., da Silva, T. H. R., da Silva Júnior, J. M., de Matos, I. E., de Carvalho, V. V., Acedo, T. S., & da Silva, A. G. (2024). Supplementation with 25-hydroxicolecalciferol increases the nutritional efficiency and carcass growth of grazing Nellore young bulls. Animal Feed Science and Technology, 318, 116138. DOI: 10.1016/j.anifeedsci.2024.116138
National Academies of Sciences, Engineering, and Medicine. (2021). Nutrient requirements of dairy cattle: Eighth revised edition. The National Academies Press. https://doi.org/10.17226/25806
Nazari Darabkhani, S., Nooriyan Soroor, M. E., & Moeini, M. M. (2024). Effect of different methods of AD3E supplementation on performance, fermentation, rumen protozoa, and blood metabolites of fattening lambs by in vivo and in vitro experiments. Animal Production Research, 13(1), 1-14. https://doi.org/10.22124/ar.2024.22957.1724
Naziroglu, M., Güler,T., & Yuce, A. (2002). Effect of vitamin E on ruminal fermentation in vitro. Journal of veterinary medicine. A, Physiology, pathology, clinical medicine,49,251– 255. doi:10.1046/j.1439-0442.2002.00418.x
Njeru, C. A., McDowell, L. R., Shireman, R. M., Wilkinson, N. S., Rojas, L. X., & Williams, S. N. (1995). Assessment of vitamin E nutritional status in yearling beef heifers. Journal of Animal Science, 73(5):1440-1448. https://doi.org/10.2527/1995.7351440x
NRC. (1985). Nutrient requirements of small ruminants: Sheep, and New World camelids. National Academies Press.
NRC. (2007). Nutrient requirements of small ruminants: Sheep, goats, cervids, and New World camelids. National Academies Press.
Oka, A., Iwaki, F., and Dohgo, T. (2004). Effects of vitamin A deficiency on growth hormone secretion and circulating insulin-like growth factor-1 concentration in Japanese Black steers. Animal Science, 78 (1): 31-36. https://doi.org/10.1017/s1357729800053819
Ottenstein, D. M., & Bartley, D. A. (1971). Improved gas chromatography separation of free acids C2-C5 in dilute solution. Analytical Chemistry, 43, 952-955. https://doi.org/10.1021/ac60302a043
Park, T., Park, S. H., & Yu, Z. (2018). Aerobic cultivation of anaerobic rumen protozoa, Entodinium caudatum and Epidinium caudatum. Journal of Microbiological Methods, 152, 186-193. https://doi.org/10.1016/j.mimet.2018.08.005
Paulo César G. Dias Junior, Isabela J. dos Santos, Fabrício L. do Nascimento, Emiro A.S. Paternina, Bárbara A. Alves, Idalmo G. Pereira, Alcinéia L.S. Ramos, Tharcilla I.R.C.
Alvarenga, & Iraides F. Furusho-Garcia. (2022). Macadamia oil and vitamin E for lambs: Performance, blood parameters, meat quality, fatty acid profile and gene expression. Animal Feed Science and Technology, 293, 115475. https://doi.org/10.1016/j.anifeedsci.2022.115475
Ramos, J.M.L., Nunes, C.L.de C., Gonçalves, H.F., Carvalho, V. V., Vasconcelos, G S F M., Acedo, T.S., Andrade,D.R., Schultz,E.B ., Oliveira,L.L., Filho,S.C.V., & Chizzotti,M.L (2024). Effect of vitamin blend supplementation on the oxidative stability and beef quality parameters of Nellore cattle. Livestock Science 286:105509
Reddy, p. & Jialal, I. (2022). Biochemistry, Fat Soluble Vitamins. NCBI Bookshelf. A service of the National Library of Medicine, National Institutes of Health. Retrieved from https://www.ncbi.nlm.nih.gov/sites/books/NBK534869/?report=printable. Assessed in Feb 13, 2026.
Santschi, D.E., Chiquette, J., Berthiaume, R., Martineau, R., Matte, J.J., Mustafa, A.F., Girard, C.L., 2005. Effects of the forage to concentrate ratio on B-vitamin concentrations in different ruminal fractions of dairy cows. Can. J. Anim. Sci. 85, 389–399. https://doi.org/10.4141/A05- 012.
Samarin, A. A., Norouzian, M. A., & Afzalzadeh, A. (2022). Effect of trace mineral source on biochemical and hematological parameters, digestibility, and performance in growing lambs. Tropical Animal Health and Production, 54(1), 40. https://doi.org/10.1007/s11250-021- 03042-1
Scapol, R. S., Baldassini, W. A., Gagaoua, M., Ramírez-Zamudio, G. D., Ladeira, M. M., Poleti, M. D., ... & Chardulo, L. A. L. (2023). Muscle proteome of crossbred cattle that received vitamin A at birth: Impacts on meat quality traits. Livestock Science, 275, 105316. https://doi.org/10.1016/j.livsci.2023.105316
Shastak, Y., Gordillo, A., & Pelletier, W. (2023). The relationship between vitamin A status and oxidative stress in animal production. Journal of Applied Animal Research, 51(1), 546-553. https://doi.org/10.1080/09712119.2023.2239319
Shirpoor, A., Ilkhanizadeh, B., Saadatian, R., Darvari, B. S., Behtaj, F., Karimipour, M., Ghaderi-Pakdel, F., & Saboori, E. (2006). Effect of Vitamin E on diabetes-induced changes in small intestine and plasma antioxidant capacity in rat. Journal of Physiology and Biochemistry, 62, 171–177 (2006). https://doi.org/10.1007/BF03168466
Shojadoost, B., Yitbarek, A., Alizadeh, M., Kulkarni, R. R., Astill, J., Boodhoo, N., & Sharif, S. (2021). Effects of vitamins A, D, E, and C on the chicken immune system, Poultry Science, 100 (4): 100930. https:/doi.org/10.1016/j.psj.2020.12.027
Silva, J. T. (2020). Effect of vitamin supplementation on the ingestive, digestive, and ruminal parameters of Nellore cattle. Master's Dissertation in Animal Science, Federal University of
Viçosa, Viçosa. Silva, É. B. R. D., Silva, J. A. R. D., Silva, W. C. D., Belo, T. S., Sousa, C. E. L., Santos, M. R. P. D., ... & Lourenço-Júnior, J. D. B. (2024). A review of the rumen microbiota and the different molecular techniques used to identify microorganisms found in the rumen fluid of ruminants. Animals, 14(10), 1448. https://doi.org/10.3390/ani14101448
Simitzis, P. E., Charismiadou, M. A., Goliomytis, M., Charalambous, A., Ntetska, I., Giamouri, E., & Deligeorgis, S. G. (2019). Antioxidant status, meat oxidative stability and quality characteristics of lambs fed with hesperidin, naringin or α-tocopheryl acetate supplemented diets. Journal of the Science of Food and Agriculture, 99(1):343-349. https://doi.org/10.1002/jsfa.9193
Skawratananond, S., McCrea, G. E., Lie, P., Buxton, M. B., Daly, S. P., Vojtkofsky, N. A., Smith, S. C., Hernandez, C. Z. M., Hindle, A., Logsdon, A. F. & Lawrence, J. J. (2025). The synergistic interplay between vitamin A, dietary fiber, and the microbiota-gut-brain axis: a potential mechanism for preventing Alzheimer’s disease. American Journal of Physiology- Gastrointestinal and Liver Physiology, 329:3, G484-G499. https://doi.org/10.1152/ajpgi.00097.2025
Śmiecińska, K., Kubiak, D., & Figger, K. (2023). The Fatty Acid Profile, and the Content of Vitamin A, Vitamin E and Cholesterol in Beef (M. longissimus lumborum) Stored Under Different Modified Atmospheres. Acta Universitatis Cinbinesis, Series E: Food Technology, 27(2). https://doi.org/10.2478/aucft-2023-0015
Song, P., Chen, X., Zhao, J., Li, Q., Li, X., Wang, Y., Wang, B., & Zhao, J. (2023a). Vitamin A injection at birth improves muscle growth in lambs. Animal Nutrition, 14, 204-212. https://doi.org/10.1016/j.aninu.2023.05.011
Song, P., Huo, G., Feng, J., Zhang, W., Li, X., & Zhao, J. (2023b). Intramuscular vitamin A injection in newborn lambs enhances antioxidant capacity and improves meat quality. Frontiers in Veterinary Science, 10, 1272874. https://doi.org/10.3389/fvets.2023.1272874
Taneera, J., Yaseen, D., Youssef, M., Khalique, A., Al Shehadat, O. S., Mohammed, A. K., Bustanji, Y., Madkour, M. I., & El-Huneidi, W. (2025). Vitamin D augments insulin secretion via calcium influx and upregulation of voltage calcium channels: Findings from INS-1 cells and human islets. Molecular and Cellular Endocrinology, 599,112472, https://doi.org/10.1016/j.mce.2025.112472
Tanumihardjo, S. A. (2011). Vitamin A: Biomarkers of nutrition for development. The American Journal of Clinical Nutrition, 94(2), 658-665. https://doi.org/10.3945/ajcn.110.005777
Tobias, D. K., Luttmann-Gibson, H., Mora, S., Danik, J., Bubes, V., Copeland, T., ... & Manson, J. E. (2023). Association of body weight with response to vitamin D supplementation and metabolism. JAMA Network Open, 6(1), e2250681. https://doi.org/10.1001/jamanetworkopen.2022.50681
Toma, F. B., Hassan, K. J., & Muhamad Salih, M. J. (2021). Role of Vitamin AD3E in Pregnancy Period and Calving of Dairy Cattle Strain Friesian. Medico-Legal Update, 21(2). https://doi.org/10.37506/mlu.v21i2.2655
Toyber, I., Kumar, R., & Jami, E. (2024). Rumen protozoa are a hub for diverse hydrogenotrophic functions. Environmental Microbiology Report, 16(4):e13298. https://doi.org//10.1111/1758-2229.13298.
Treacher, R. J. (1978). Dietary protein levels and blood composition of dairy cattle. BSAP Occasional Publication, 1, 133-142. https://doi.org/ 10.1017/S0263967X00000148
Upadhaya, S. D., Chung, T. K., Jung, Y. J., & Kim, I. H. (2023). Efficacy of dietary vitamin D3 and 25(OH)D3 on reproductive capacities, growth performance, immunity and bone development in pigs. British Journal of Nutrition, 130(8), 1298-1307. https://doi.org/10.1017/S0007114523000442
Velykyĭ, M, M., Apukhovs'ka, L. I., Vasylevs'ka, V. M., Lotots'ka, OIu., Besusiak, A. I., & Khomenko, A. V. (2010). Features of cholecalciferol hydroxylation in the liver of rats in conditions of D-hypervitaminosis and activity of alpha-tocopherol. The Ukrainian Biochemical Journal, 82(2):67-74. https://pubmed.ncbi.nlm.nih.gov/20684247
Verdurico, L. C. (2012). Evaluation of Mediterranean buffalo cows during the transition period and early lactation and young calves until weaning. Universidade de Sao Paulo, Agencia USP de Gestao da Informacao Academica (AGUIA). https://doi.org/10.11606/D.10.2010.tde- 11012012-163851
Vijayalakshmi, B., Sesikeran, B., Udaykumar, P., Kalyanasundaram, S., & Raghunath, M. (2005). Effects of vitamin restriction and supplementation on rat intestinal epithelial cell apoptosis. Free Radical Biology and Medicine, 38 (2):1614-1624. https://doi.org/10.1016/j.freeradbiomed.2005.02.029
Voia, S., Alhorani, I., Drinceanu, D., Padeanu, I., Sauer, M., Gavojdian, D., & Lunca, M. (2011). Influence of nutritive supplement on the stimulation of ruminal microorganism’s activity in
fattening lambs reared in a semi-intensive system. Animal Science and Biotechnologies, 44 (1): 145-148. CABI code: 20133359118
Wang, D., Dal Jang, Y., Rentfrow, G. K., Azain, M. J., & Lindemann, M. D. (2022a). Effects of dietary vitamin E and fat supplementation in growing-finishing swine fed to a heavy slaughter weight of 150 kg: I. Growth performance, lean growth, organ size, carcass characteristics, primal cuts, and pork quality. Journal of Animal Science, 100, skac081. https://doi.org/10.1093/jas/skac081
Wang, J., Wu, S., Zhang, Y., Yang, J., & Hu, Z. (2022b). Gut microbiota and calcium balance. Frontiers Microbiology, 13: 1033933. https://doi.org/10.3389/fmicb.2022.1033933
Wei, C., Lin, S. X., Wu, J. L., Zhao, G. Y., Zhang, T. T., & Zheng, W. S. (2015). Effects of supplementing vitamin E on in vitro rumen gas production, volatile fatty acid production, dry matter disappearance rate, and utilizable crude protein. Czech Journal of Animal Science, 60(8), 335-341. https://doi.org/10.17221/8402-CJAS
Witariadi, N. M., Kusumawati, N. N. C., Made, N., & Sukmawati, S. (2022). Effect of substitution urea fertilizer with cow manure fertilizer on the productivity of Asystasia gangetica (L.) Subsp. Micrantha. International Journal of Fauna and Biological Studies, 9(6), 49-52. https://doi.org/10.22271/23940522.2022.v9.i6a.944
Xiao, J., Khan, M. Z., Ma, Y., Alugongo, G. M., Ma, J., Chen, T., ... & Cao, Z. (2021). The antioxidant properties of selenium and vitamin E; their role in periparturient dairy cattle health regulation. Antioxidants, 10(10), 1555. https://doi.org/10.3390/antiox10101555
Yoon, J. H., Ingale, S. L., Kim, J. S., Kim, K. H., Lohakare, J., Park, Y. K., Park, J. C., Kwon, I. K., & Chae, B. J. (2013). Effects of dietary supplementation with antimicrobial peptide P5 on growth performance, apparent total tract digestibility, faecal and intestinal microflora and intestinal morphology of weanling pigs. Journal of the Science of Food and Agriculture, 93, 587–592. https://doi.org/10.1002/jsfa.5840
Yu, B., Chen, j., Wang, Y., Zhou, J., Wang, H., Li, H., Cai, T., Huang, R., Zhou, Y., & Ma, J. (2024). Vitamin A influences the incretin hormone profiles by activating the retinoic acid receptor β. Journal of Diabetes and its Complications, 38 (8), 108806, https://doi.org/10.1016/j.jdiacomp.2024.108806
Zayed, Z., Estuty, N., & Abdelatti, S. (2021). Effects of Dietary Vitamin E Supplementation on Haematological Parameters, Serum Vitamin E and Lipid Oxidation of Mahali Goats In Libya. Assiut Veterinary Medical Journal, 67(170), 51– 58. https://doi.org/10.21608/avmj.2021.98640.1025
با محصولات دامی بیشتر آشنا شوید:
با محصول مکمل بافری بهدام رشد (متوسط تولید) و ( مخصوص کارخانجات) بیشتر آشنا شوید
با محصول مکمل انتظار زایش بیشتر آشنا شوید
با محصول مکمل دامی بیشتر آشنا شوید
با محصول مکمل آنیونی بیشتر آشنا شوید
با محصول کنسانتره 4 درصد گاو شیری (پرتولید) بیشتر آشنا شوید
با محصول کنسانتره 4 درصد گاو شیری (متوسط تولید و پرواری) بیشتر آشنا شوید
با محصول مکمل بافری بهدام رشد (پرتولید) بیشتر آشنا شوید
با محصول کنسانتره 5 درصد گاو شیری (ممتاز) بیشتر آشنا شوید
با محصول کنسانتره 3/5 درصد گوسفندی (میش داشتی آبستن سبک) بیشتر آشنا شوید
با محصول کنسانتره 3/5 درصد گوسفندی (آبستن سنگین) بیشتر آشنا شوید
با محصول کنسانتره 3/5 درصد گوسفندی (پرواری) بیشتر آشنا شوید
با محصول کنسانتره 3/5 درصد فلاشینگ بیشتر آشنا شوید



